|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Альтернативный путь активацииРаспознавание бактерий и других активирующих поверхностей их
молекул
опсонизация дегрануляция тучных клеток
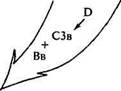
регуляция Т-системы выработка ПГ включение альтернативного пути образование растворимых комплексов Хемотаксис ЛИЗИС КЛЕТОК Рис. 4.11. Система комплемента цитоза и аллергических реакциях. Так, пептиды СЗа и С5а обладают свойствами анафилотоксина. Присоединяясь к тучным клеткам и базофилам, они индуцируют выделение гистамина. Связываясь с тромбоцитами, СЗа вызывает секрецию серотонина. Ана-филотоксическая активность СЗа и С5а легко разрушается под воздействием карбоксипептидазы В, которая отщепляет от этих пептидов аргинин. Образующиеся продукты приобретают свойства хемоаттрактантов в отношении полиморфноядерных клеток, эозинофилов и моноцитов. Другой пептид — СЗв является сильным опсонином для полиморфноядерных клеток и макрофагов. Рецепторы к этому пептиду обнаружены и на других клетках: В-лимфо-цитах и моноцитах. Наличие на В-лимфоцитах рецепторов для СЗ используется в качестве одного из основных маркеров этой популяции. Взаимодействие СЗ и его субкомпонентов (СЗв, СЗс, C3d) с В-лимфоцитами играет определенную роль в индукции специ-
12 — 3389 фического иммунного ответа и в регенерации В-клеток памяти. Установлено также участие СЗ в продукции антител к Т-зависи-мым антигенам и во взаимодействии Т- и В-клеток, а также макрофагов, Т- и В-клеток. Известно, что С5 участвует в антителоза-висимой цитотоксичности лимфоцитов, осуществляя на поверхности лимфоцитов сборку комплементарного мембранолитичес-кого комплекса. Связанная с мембраной макрофагов С1 компонента играет роль в фиксации комплекса антиген—антитело. Система комплемента имеет большое значение для диссоциации и элиминации иммунных комплексов (ИК). Такое участие обеспечивается связыванием СЗв, который, соединяясь с антителом, снижает способность связывания антигена с Fab-фрагментом. В данном процессе задействован также С4в. Эти факторы комплемента не только препятствуют образованию иммунных комплексов, но также участвуют в разрушении уже сформировавшихся. Уменьшение или увеличение содержания комплемента наблюдается при многих заболеваниях (воспалительные процессы, аутоиммунные болезни, опухоли). У собак породы британский спаниель встречается врожденный дефицит СЗ фрагмента комплемента. Дефицит СЗ компонента наследуется по аутосомно-рецессивному типу и клинически проявляется часто повторяющимися бактериальными инфекциями у гомозиготных индивидуумов. В результате дефицита комплемента, уровень которого составляет лишь 10 % от нормального, снижаются опсонизация, хемотаксис и иммуноприлипание, что проявляется повышенной чувствительностью к инфекциям. Гуморальный и клеточный иммунитет у пораженных британских спаниелей остается в норме. Одним из основных действий И К является активация плазменных компонентов системы комплемента и иммунокомпетентных клеток. Комплемент играет важную роль в выведении ИК из организма, поэтому способность ИК взаимодействовать с компонентами классического либо альтернативного пути системы комплемента в конечном итоге определяет характер воспаления и тканевого повреждения в организме. Источником комплемента являются клетки нескольких типов, включая тканевые макрофаги, гепатоциты, кератиноциты, клетки слизистой оболочки толстой кишки, эндотелиальные клетки, по-лиморфноядерные лейкоциты. Печень является источником более чем 90 % плазменных белков, а макрофаги — основным источником тканевого комплемента, особенно в условиях воспаления. Интенсивность биосинтеза этих компонентов может существенно меняться в зависимости от количества и типа ИК, находящихся в циркуляции. На синтез компонентов комплемента помимо ИК влияют системно действующие гормоны, интерлейкины и биологически активные соединения. Система комплемента играет важную роль в процессе растворения ИК. Взаимодействие циркулирующего иммунного комплекса (ЦИК) с системой комплемента обеспечивает растворение крупных нерастворимых ИК до мелких. В опытах in vitro показано, что нерастворимые ИК становятся растворимыми при добавлении свежей сыворотки при 37 °С. Инициируемая комплементом солюбилизация ИК является следствием связывания этих комплексов с СЗв таким образом, что процесс солюбилизации ИК является СЗ-зависимыми. Частичное растворение ИК происходит и в СЗ, С4-дефицитной сыворотке, но не в сыворотке при поврежденном альтернативном пути активации комплемента. Компоненты альтернативного пути активации комплемента пропердин и фактор D наряду с факторами В, СЗ и Mg2+ также играют важную роль в растворении ИК. Классический путь сам по себе не обеспечивает растворения, однако его активизация приводит к значительному повышению в крови количества СЗв и увеличению вероятности связывания с комплексами антиген — антитело. Таким образом, компоненты классического пути повышают эффективность активации компонентов альтернативного пути в процессе растворения ИК. Важнейшим аспектом взаимодействия ЦИК и системы комплемента является изменение физико-химических свойств самого комплекса в процессе присоединения к нему различных компонентов комплемента, что приводит к увеличению степени дисперсности и уменьшению агрегации комплексов. Взаимодействие ИК и системы комплемента является ключевым моментом в судьбе ЦИК, поскольку помимо активации системы комплемента это взаимодействие ведет к возможности присоединения ИК через Fc- и С-рецепторы к большинству иммунокомпетентных клеток, что влияет на Т—В-взаимодействия, изменяет фагоцитарную активность клеток. Активация фагоцитарной системы приводит либо к удалению комплекса из кровотока, либо способствует длительной циркуляции, дальнейшему отложению ИК в органах и тканях и развитию васкулитов. Взаимодействие ИК и системы комплемента приводит к двум основным следствиям: образованию фрагментов компонентов комплемента, обладающих разносторонней биологической активностью, и ингибированию преципитации ИК при активации по классическому пути либо растворению сформировавшихся уже комп-цексов при определяющем участии компонентов альтернативного пути активации. В нормальной сыворотке крови компоненты классического пути поддерживают ИК в растворимом состоянии в течение времени, достаточного для их элиминации мононуклеар-пыми фагоцитами. Компоненты альтернативного пути не спо-гобны ингибировать преципитацию ИК, но могут солюбилизиро-кать агрегаты антиген — антитело. Взаимодействие ЦИК с системой комплемента не только приводит к связыванию ИК с рети- i>* 179
Растворенные ИК не могут фиксировать комплемент и почти полностью лишены сродства к поверхностным рецепторам различных клеток. Комплемент ускоряет клиренс растворимых ИК, осуществляемый фагоцитами. На растворение ИК существенно влияет свойство комплекса фиксировать комплемент. ИК с некоторым избытком антигена под влиянием свежей сыворотки растворяются не полностью, а ИК с большим избытком антигена не растворяются компонентами ни альтернативного, ни классического пути активации системы комплемента; ИК с избытком антигена растворяются компонентами только альтернативного пути (GaninG et al., 1983). ИК, образованные вне сосудистых пространств, удаляются значительно медленнее и могут провоцировать местные воспаления. В заключение можно сказать, что аномалии в системе комплемента способствуют развитию иммунокомплексных болезней. Дефицит в системе комплемента приводит к нарушению связи ИК — комплемент — дендритная клетка лимфатического узла, что, в свою очередь, влияет на иммунный ответ в целом. Пропердин (лат. perdere — разрушать) — белок, с помощью которого обнаружен альтернативный механизм активации комплемента. Он представляет собой гамма-глобулин с молекулярной массой 220000 и состоит из четырех практически идентичных субъединиц, соединенных друг с другом нековалентными связями. Его концентрация в сыворотке составляет около 25 мкг/мл. Пропердин существует в двух формах: нативной и активированной, различающихся между собой, по всей видимости, небольшими конформационными изменениями. Нативный пропердин может связываться с образовавшей комплекс СЗ/С5-конвертазой альтернативного механизма (СЗвВв), но не с одиночными молекулами СЗв. Его роль заключается в уменьшении скорости распада конвертазы и тем самым усилении активации по альтернативному механизму. Пропердин действует, таким образом, не сам по себе, а совместно с другими факторами, содержащимися в крови животных, в том числе и с комплементом. Сама же система комплемента состоит из трех основных частей: пропердина, ионов Mg+2, комплемента. Активация пропердина осуществляется СЗ-компонентом комплемента. Пропердиновая система обладает антибактериальным действием в отношении многих патогенных и условно патогенных микроорганизмов. Под действием пропердина инактиви-руются вирусы герпеса и гриппа. Показатель уровня пропердина в крови в определенной мере отражает чувствительность животных к инфекции. Установлено, что происходит снижение содержания пропердина при туберкулезе, стрептококковой инфекции, ионизирующем облучении. Изъятие из сыворотки крови пропердина резко снижает ее нейтрализующую активность. Полная инактивация пропердина происходит при нагревании до 60 "С в течение 30 мин. ЛИЗОЦИМ Лизоцим — фермент, относящийся к классу гидролаз, избирательно гидролизующий гликозидные связи в муреине — сложном биополимере, из которого построены стенки бактерий. Молекулярная масса лизоцима 14000...15 000. Это стойкий белок, не теряющий литической способности при нагревании до 100 "С. Способность лизоцима лизировать микроорганизмы столь высока, что это свойство сохраняется в разведении 1: 1 000 000. Его молекула состоит из 129 аминокислотных остатков, представлена одной полипептидной цепью, содержащей 8 половинок цистина, попарное соединение которых образует четыре дисульфидные связи. Они замыкают спиральные участки полипептидной цепи лизоцима. Молекула лизоцима окружена гидрофобными группами боковых цепей остатков аминокислот. Главная роль в образовании активного центра принадлежит, по-видимому, триптофану. Ферментативная активность лизоцима проявляется в гидролизе 1,4-гликозидной связи полиаминосахаров клеточной стенки преимущественно грамположительных микроорганизмов. Абсорбируясь мукопептидом клеточной стенки, лизоцим расщепляет его с освобождением N-ацетилмурамовой кислоты и N-ацетилглю-козамина. Искажение структуры субстрата, поляризация гликозид-ной связи, образование водородной связи с кислородом последней приводят совместно к разрыву глюкозидной связи, а окружающая вода завершает акт гидролиза. Скорость реакции расщепления субстрата у разных лизоцимов различна, что, вероятно, связано с различием первичной структуры разных лизоцимов. Лизоцим обнаружен в различных тканях и секретах: в сыворотке крови, слезах, слюне, молоке. Его максимальное количество содержится в лейкоцитах, затем в слюне и слезах, минимальное — в сыворотке крови. Почки денатурируют и разрушают плазменный лизоцим. В плазму крови лизоцим поступает при распаде лейкоцитов и тканей. Концентрация его зависит от соотношения между основными продуцентами — нейтрофилами и моноцитами и функции почек. Макрофаги высвобождают лизоцим постоянно, гранулоциты — только при дегрануляции, поэтому сывороточный лизоцим может служить индикатором макрофагальной функции организма. Основываясь на антибактериальных свойствах лизо-
ИНТЕРФЕРОНЫ Интерфероны — антивирусные агенты. Существует по крайней мере 14 альфа-интерферонов, которые продуцируются лимфоцитами, а бета-интерферон—фибробластами. При вирусной инфекции клетки синтезируют интерферон и секретируют его в межклеточное пространство, где он связывается с рецепторами соседних незараженных клеток. Связанный с клеткой интерферон дерепрессирует по меньшей мере два гена. Начинается синтез двух ферментов: первый — протеинкиназа значительно снижает в конечном итоге трансляцию мРНК; второй — катализирует образование короткого полимера аде-ниловой кислоты, активирующего латентную эндонуклеазу, что приводит к деградации мРНК как вируса, так и хозяина. В целом конечный результат действия интерферона заключается в образовании барьера из неинфицированных клеток вокруг очага вирусной инфекции, чтобы ограничить ее распространение. Интерфероны играют большую роль в борьбе с вирусами, но не в предотвращении вирусных инфекций. Система нормальных киллеров. К лимфоидным клеткам, способным оказывать цитотоксическое действие без сенсибилизации, относятся NK-клетки (естественные киллеры), которые в отличие от К-клеток могут проявлять цитотоксическое действие и в отсутствие специфических антител. Биологическое действие NK-клетки связано с контролем раннего опухолевого развития. NK-клетки обладают цитотоксической активностью по отношению к различным опухолевым клеткам, а также к клеткам, инфицированным вирусными или микробными агентами. Благодаря этому NK могут играть важную роль в устойчивости организма ко многим заболеваниям. при этом соединяется своей антигенной детерминантой, антитело — своим активным центром. При избытке антигенов или антител образуются растворимые комплексы, при эквивалентном соотношении — нерастворимый преципитат. Антиген, как правило, крупнее молекулы антитела, поэтому последняя может распознавать только отдельные участки антигена, которые называют детерминантами. Большинство антигенов имеет на поверхности целый набор различных антигенных детерминант, каждая из которых стимулирует иммунный ответ. Не все они одинаковы по активности: одни более иммуногенны и реакция на них доминирует в общем ответе. Даже одиночная детерминанта активирует, как правило, разные клоны клеток с поверхностными рецепторами (антителами), которые обладают разным сродством к данной детерминанте. Следовательно, иммунный ответ на большинство антигенов является политональным. Вместе с тем образовавшиеся антитела могут вступать в реакцию не только с гомологичным антигеном, но и с родственными ему гетероло-гичными антигенами. Реакции неспецифического взаимодействия антител сыворотки крови с антигенами проявляются в следующих формах: агглютинация — склеивание антигенных частиц между собой; преципитация — агрегация частиц с образованием нерастворимых комплексов; лизис — растворение клеток под влиянием антител в присутствии комплемента; цитотоксичность — гибель клеток под влиянием антител — цитотоксинов; нейтрализация — обезвреживание токсинов белковой природы; опсонизацш — усиление фагоцитарной активности нейтрофилов и макрофагов под влиянием антител или комплемента. Обычный иммунный ответ выявляется через несколько суток после связывания антигена с В-лимфоцитом. Он представляет собой интегральную реакцию организма на антиген вследствие сложных взаимодействий между клетками разных типов. Дата добавления: 2015-12-15 | Просмотры: 759 | Нарушение авторских прав |
 кулоэндотелиоцитами, но и обеспечивает переход нерастворимых ИК в растворимые или их полный распад. В процессе растворения ИК определяющая роль принадлежит компонентам альтернативного пути. Классический путь сам по себе не обеспечивает растворения ИК, но способствует повышению эффективности этого процесса за счет продукции СЗ-конвертазы.
кулоэндотелиоцитами, но и обеспечивает переход нерастворимых ИК в растворимые или их полный распад. В процессе растворения ИК определяющая роль принадлежит компонентам альтернативного пути. Классический путь сам по себе не обеспечивает растворения ИК, но способствует повышению эффективности этого процесса за счет продукции СЗ-конвертазы. цима, большинство исследователей склонно рассматривать его как фактор неспецифического иммунитета. Кроме основного антибактериального действия лизоцим стимулирует естественную резистентность организма животного, что играет большую роль в предупреждении заболеваний и в благоприятном исходе инфекционного процесса.
цима, большинство исследователей склонно рассматривать его как фактор неспецифического иммунитета. Кроме основного антибактериального действия лизоцим стимулирует естественную резистентность организма животного, что играет большую роль в предупреждении заболеваний и в благоприятном исходе инфекционного процесса.