|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Поперечнопосмугована м'язова тканинаСкелетна (соматична) м'язова тканина — це робоча мускулатура апарату руху, глотки, гортані, верхньої половини стравоходу, язика, жуйних м'язів. Джерелом розвитку цієї тканини є матеріал міотпомів сегментованої мезодерми, а поперечнопосмугована тканина внутріш-ніх органів розвивається із спланхнотому. На ранній стадії розвитку міотоми складаються із щільно розміщених м'язових клітин — міо-бласшів, цю стадію гістогенезу називають міобластичною. Міобласти диференціюються у двох напрямах, одні здатні злива-тися і утворювати симпласти — м'язові трубочки, які далі формують дефінітивні структури — міосимпласти. У результаті другої лінії дифе-ренціації розвиваються клітинні структури — міосателітоцити. На першій міобластичній стадії гістогенезу (рис. 62 А, Б) цито-плазма міобластів має тонковолокнисту будову, що характеризує розвиток скорочувальних білків. Уже на цій стадії міобласти здатні до скорочення. Ядра їх великі, овальні з невеликим вмістом гетеро-хроматину. Вони здатні до інтенсивнішого поділу, ніж сама клітина, в результаті чого міобласти стають багатоядерними, витягуються у довжину і набувають форму волокна — симпласта. Міосимпласти можуть утворюватися і шляхом злиття міобластів — це друга стадія гістогенезу — м 'язових трубочок. В них міофібрили розташовуються під сарколемою, а ядра в центрі. М'зові трубочки здатні розщеплюва-тися вздовж і формувати м'язові волокна, в яких різко збільшується кількість міофібрил. Численні ядра зміщуються на периферію під плазмолему, волокна набувають посмугованості. Це третя стадія гіс-тогенезу — м 'язових волокон. До м'язових волокон підростає сполучна тканина з кровоносними судинами, нервами, які диференціюються в нервові закінчення.
Розділ 3 Загальна гістологія
Рис. 62. Етапи ембріогенезу поперечнопосмугованої м'язової тканини: а-клітина соміта (1-міотом; 2-дерматом); б-міобласти; в-міосимпласти; г-про-міотуб; д-м'язова трубочка; е-незріле м'язове волокно; є-зріле м'язове волок-. но; ж-сполучнотканинна клітина. Наве^ені дані про гістогенез допоможуть зрозуміти будову сома-тичної м'язової тканини та ті складні зміни, які у ній відбуваються під час тренування, фізичних навантажень та при фізіологічній регенера-ції і патології. У диференційованої скелетної м'язової тканини відсутня клітин-на будова. Основною структурною і функціональною одиницею її є м'язове волокно. Довжина його часто збігається з довжиною м'яза, до складу якого воно входить. Волокно покрите сарколемою (від гр. 5агкоз — м'ясо), яка складається із двох мембран. Зовнішня базаль-
В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, ггстологія, ембріологія на мембрана стикується із ретикулярними та тонкими колагенови-ми волокнами сполучної тканини, що її оточує. Внутргшня мембрана аналогічна плазмолемі усіх тканинних клітин і бере участь у про-веденні імпульсів скорочення волокна. Плазмолема утворює систе-му вузеньких трубочок, які пронизують все волокно і зливаються із сарколемою протилежного боку. Таким чином, вся саркоплазма про-низана системою поперечних трубок, Т-системою. Цитоплазма має спеціальну назву — саркоплазма, структурними компонентами якої є міофібрили (скорочувальний апарат волокна), органели, включен-ня, гіалоплазма. Численні ядра м'язового волокна розміщуються на периферії під сарколемою. У гіалоплазмі волокна локалізуються мі-тохондрії (саркосоми), які скупчуються між мікрофібрилами та на-вколо численних ядер. Це ті зони волокна, які потребують значної кількості АТФ, через це стає зрозумілою висока метаболічна актив-ність скелетних м'язів. Найбільш інтенсивного розвитку досягає агранулярна ендоплаз-матична сітка (саркоплазматичний ретикулум). її мембранні елемен-ти розміщуються вподовж саркомерів (частинки, з яких складається волокно й залежить його довжина). Саркоплазматичному ретикулу-му притаманна специфічна функція нагромадження іонів кальцію, необхідних при скороченні та розслабленні м'язового волокна. Інші органели (гранулярна ендоплазматична сітка, комплекс Гольджі) розвинені менше і локалізуються біля ядер, розміщених на периферії під сарколемою. Ядра можуть розміщуватися у вигляді ланцюжка один за одним, що є в результаті амітотичного поділу — показник реактивного стану м'язового волокна. Кількість ядер коливається від десятка до декіль-кох сотень, вони мають овальну форму. Гетерохроматин у вигляді ве-ликих гранул знаходиться у порівняно світлій нуклеоплазмі. Поміж міофібрил локалізується значна кількість гранул глікогену (трофічного) включення — матеріалу для синтезу АТФ. У цитоплазмі м'язового волокна містяться дихальні ферменти, білок, міоглобін — ана-лог гемоглобіну еритроцитів, який також здатний зв'язувати та віддава-ти кисень. Міоглобін забарвлює м'язові волокна у червоний колір. В залежності від вмісту саркоплазми (а, отже, і міоглобіну), товщи-ни і ферментного складу м'язові волокна поділяють на червоні та білі. Червоні волокна у саркоплазмі містять велику кількість міоглобіну, Розділ 3 Загальна гістологія численні мітохондрії, багаті на цитохроми, міофібрили у них мають незначну товщину. М'язи, у яких переважають червоні волокна, здатні до тривалішої безперервної активності, оскільки їх саркоплазма добре забезпечує енергетичні потреби. Білі волокна містять у саркоплазмі менше міоглобіну та мітохон-дрій, у них більша товщина міофібрил. Білі м'язові волокна здатні ско-рочуватися швидше, ніж червоні, али вони порівняно швидко втомлю-ються, тому що не забезпечують структури саркоплазми достатньою кількістю енергії. Виходячи з цього, можна порівняти робочу силу і втомлюваність грудних м'язів птахів, що літають і тих, що не здатні до польоту; м'язи треновані і нетреновані у молодої та старої особини. Найбільшу питому вагу у саркоплазмі становлять міофібрили. Вони розміщуються вздовж м'язового волокна і їх довжина збігається з довжиною волокна, діаметр міофібрил — 1-2 мкм. Вони мають харак-терну поперечну посмугованість (чергування світлих і темних смуг, дисків), що зумовлено особливістю їхньої структури і, в зв'язку з цим, різними оптичними властивостями. Усі темні й всі світлі диски міо-фібрил в одному волокні містяться на одному рівні, через що волокно має поперечну посмугованість. Повздовжня орієнтація міофібрил на-дає м'язовому волокну поздовжню смугастість. У поляризованому світлі темні смуги (диски) мають подвійне про-менезаломлення — анізотропію, через що їх називають анізотропними (дисками) А. Світлі смуги (диски) є однопроменезаломлюючими, через що їх називають ізотропними — I. Всередині кожного І-диска про-ходить темна зона — лінія 2 (телофрагма). У центрі А-диска спосте-рігається свгтлгша зона-лінія Н, або смужка Гензена, поеередині якої розміщується тонка, темна лінія М (мезофрагма). Міофібрили можна одержати, розщепивши м'язове волокно. Структурною одиницею міофібрил є саркомер. Це ділянка міофі-брили між двома телофрагмами. В міофібрилі саркомери розміщують-ся один за одним і в зоні Т-телофрагми, багатої на глікозаміноглікани, міофібрили можуть при мацерації розпадатися на окремі саркомери. Саркомер — це частина міофібрили, що складається із Т-телофрагми — лінії 2 (для двох сусідніх саркомерів), половин І-диска та А-диска половини зони Н, мезофрагми-М, половини зони Н, половин А- і І-дисків, лінії 2-телофрагми. В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія Структурну формулу саркомера можна записати так: Т|2| + 1/2 + А/2 + Н/2 + М +Н/2 + А/2 +1/2 +Т|2| Саркомери — це елементарні скоротливі одиниці поперечно-сму-гастих м'язів, які скорочуються завдяки здатності зменшувати свою до-вжину в два рази. Електронномікроскопічними, гістохімічними, біохі-мічними дослідженнями вдалося встановити функціональну морфоло-гію саркомера. Були ідентифіковані поздовжні нитки саркомера-міофі-ламенти або мікрофіламенти двох типів. А-диск складається із товстих міофіламентів (діаметр — 10-12 нм, довжина— 1,5-2 мкм), І-диск— із тонких (діаметр — 5-7 нм, довжина — 1-1,3 мкм) міофіламентів. Мате-ріалом, з якого складаються товсті міофіламенти, є білок міозін, а тон-кі — актин, тропоміозин-В і тропін. Кількісне відношення міозинових і актинових міофіламентів в одній міофібрилі 1:2, тобто на один міозино-вий міофіламент припадає два актинових. Актинові й міозинові міофі-ламенти контактують один з одним не кінець в кінець, а переміщуються (ковзають) по відношенню один до одного і в А-диску утворюють зону перекриття. Частину А-диска, яка складається лише з міозинових міо-філаментів, називають Н-лінією і, порівняно із зоною перекриття, вони світліші. При скороченні саркомера актинові міофіламенти ще далі про-никають у проміжки між міозиновими, а при повному скороченні їхні вільні кінці майже збігаються у середині саркомера (рис. 63). Оскільки довжина таких філаментів залишається незмінною, вони, просочуючись між товстими філаментами, тягнуть за собою телофраг-ми (2-пластини), до яких прикріплені, тим самим зближуючи кінці усіх саркомерів. У повністю скороченому саркомері Н-зона, а також І-диски майже зникають і весь саркомер перетворюється на зону пе-рекриття. М-лінія (мезофрагма) — це місце з'єднання товстих міози-нових міофіламентів в анізотропному А-диску, 2-лінія (телофрагма) проходить через усю товщину саркомера, а зона прикріплення тонких актинових міофіламентів має зигзагоподібний контур. 2-лінія склада-ється із 2-філаментів, до складу яких входять білки — тропоміозин-В та ь-актин, 2-філаменти формують решітку, до якої з обох боків при-кріплюються тонкі актинові міофіламенти І-дисків двох сусідніх сар-комерів. Таким чином, телофрагми (2-лінії) та мезофрагми (М-лінії) є опорним апаратом саркомерів. Отже, під час скорочення м'язового Розділ 3 Загальна гістологія волокна його скоротливий апарат зазнає таких змін. Зменшується довжина саркомера, оскільки актинові міофіламенти І-диску просу-ваються (ковзають) між міозиновими А-диску, зсуваючись до Н-лінії (мезофрагми) А-диску. Збільшуються зони перекриття, формуючи бокові з'єднання (місточки) між актиновими та міозиновими міофі-ламентами; скорочується Н-лінія (мезофрагма); зближуються 2-лінії (телофрагми). Чим сильніше скорочується міофібрила, тим глибше актинові міофіламенти заходять у проміжки між міозиновими, зони перекриття розширяються за рахунок звуження Н-ліній.
1/21-диск А-диск 1/21-диск
Рис. 63. Саркомер поперечнопосмугованого м'язового волокна в розслабленому (I) та скороченому (II) станах: 1-тонкі міофіламенти; 2-товсті міофламенти; 3-зони перекриття. Для розуміння механізму скорочення міофібрили необхідно зга-дати про наявність спеціалізованого саркоплазматичного ретикулуму В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія саркоплазми волокна та утворення плазмолемою системи поперечних канальців Т-трубочок. Зони контакту системи Т-трубочок і термі-нальних цистерн саркоплазматичного ретикулуму називають трі-адами. Т-трубки проходять на рівні 2-ліній (телофрагми) через все волокно і контактують з цистернами саркоплазматичної сітки з проти-лежного боку. Таким чином, Т-трубочки локалізуються на межі двох саркомерів і контактують з терміналями саркоплазматичної сітки обох саркомерів, утворюючи тріади. Ці структури відіграють основну роль в деполяризації (поширенні імпульсу) та акумуляції іонів кальцію. Плазмолема м 'язового волокна, так як і неврилема нервових воло-кон, електрично поляризована. Внутрішня поверхня плазмолеми, роз-слабленого м'язового волокна, має негативний потещіал, а зовнішня-позитивний. При скороченні м'язових волокон хвиля деполяризації по нервовому волокну через нервове закінчення досягає плазмолеми м'я-зового волокна і викликає її місцеву деполяризацію. Через систему Т-трубочок, яка зв'язана з плазмолемою та тріадою, хвиля деполяризації впливає на проникність мембран саркоплазматичного ретикулуму, що призводить до звільнення акумульованих іонів кальцію з її поверхні у саркоплазму. В присутності іонів кальцію активізується розщеплення АТФ, що необхідно для утворення актоміозинового комплексу та ков-зання актинових міофіламентів по відношенню до міозинових. Це ви-кликає скорочення кожного саркомера, а звідси міофібрил та м'язових волокон у цілому (рис. 64). Важливе місце у цьому процесі належить молекулам міофіламен-тів-міозину. Вони складаються із головки та довгого хвостика. При гідролізі АТФ, чому сприяє АТФ-на активність голівок молекул міо-зину, вони утворюють зв'язки (містки) з певними частинами молекул міофіламентів-актину. Актинові міофіламенти зближаються до цен-тру саркомеру, 2-лінії (телофрагми) зближуються, збільшуються зони перекриття, зменшуються Н-зони (мезофрагма) анізотропних дисків міофібрил. Потім з участю АТФ актоміозинові зв'язки руйнуються, а міозинові головки приєднуються до сусідніх ділянок актинових міо-філаментів, що сприяє дальшому просуванню міофіламентів один до одного. При зменшенні концентрації іонів кальцію (вони трансфор-муються на мембрани саркоплазматичного ретикулуму) скорочення м'язового волокна припиняється. Для цього також необхідна енергія АТФ. Отже, при розслабленні так, як і при скороченні м'язового во- Розділ 3 Загальна гістологія локна витрачається АТФ, джерелом якої в саркоплазмі є глікоген, глю-коза та жирні кислоти.
1-саркоплазматичний ретикулум; 2-термінальні цистерни саркоплазматич-ного ретикулуму; З-Т-трубки; 4-тріади; 5-сарколема; 6-міофібрили; 7-А-диск; 8-І-диск; 9-2-лінія; 10-мітохондрії. Кінці м'язових волокон міцно фіксуються до сухожилків або сухо-жильних прошарків, що розміщуються між ними. Сарколема утворює пальцеподібні вирости, між якими знаходяться колагенові волокна сполучної тканини, які кріплять м'язові волокна до кісток. Цей зв'язок настільки міцний, що при навантаженні, яке здатне розірвати м'язи або сухожилки, структура залишається цілою. Тонкі прошарки рихлої сполучної тканини між м'язовими волокнами називаються ендомізієм, ретикулярні і колагенові волокна його переплітаються з волокнами В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія сарколеми (зовнішній сполучно-тканинний шар). В ендомізії локалі-зуються гемокапіляри та структури нервової тканини. Комплекс во-локна з оточуючими його елементами є структурною і функціональ-ною одиницею скелетного м'яза. М'язові волокна об'єднуються у пуч-ки, між якими наявні товщі прошарки рихлої сполучної тканини, яка носить назву перимізій. Сполучна тканина, що покриває м'яз у цілому, як орган, називається епімізієм. Поперечнопосмугована м'язова тканина здатна до активної ре-генерації. Репаративна регенерація відбувається на фоні відмирання старих структур і утворення нових. Як і при нормальному гістогенезі, регенерація відбувається у три фази: міобластичну; м'язових трубочок; формування м'язового волокна. Дата добавления: 2015-12-15 | Просмотры: 866 | Нарушение авторских прав |



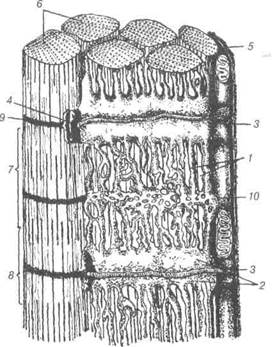
 Рис. 64. Схема будови ділянки м'язового волокна:
Рис. 64. Схема будови ділянки м'язового волокна: