|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
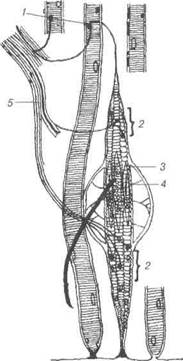
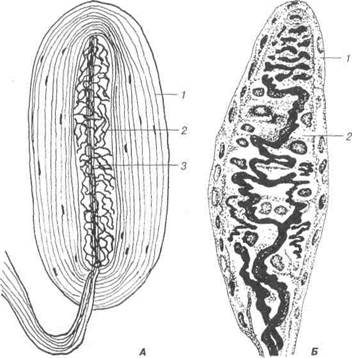
Нервові волокнаНервові волокна — це провідники нервових імпульсів. Складають-ся вони із відростка нейрону — осьового циліндра і оболонки, що його вкриває (нейролемоцит, швановська клітина) — структура олігоден-дроглії. Залежно від будови оболонки їх поділяють на дві основні гру-пи — мієлінові та безмієлінові. Безмієлінові нервові волокна (рис. 74) є типовими для вегетатив-ного відділу нервової системи, порівняно просто побудовані і спрямо-вуються, переважно, до внутрішніх органів. Нейролемоцити щільно прилягають один до одного, утворюючи безперервний тяж на поверхні нервового волокна. Складаються безмієлінові волокна з осьового ци-ліндра, нейролеми і базальної мембрани. Прогинаючи оболонку ней-ролемоцитів, осьовий циліндр глибоко занурюється у цей тяж клітин, які, ніби муфта, огортають нервовий відросток. Входячи до лемоцита, осьові циліндри втягують за собою мембрану нейролемоцита і стають немов підвішеними на подвійній складці мембрани, яку називають ме-заксоном. В безмієліновому нервовому волокні міститься від 7 до 10 осьових циліндрів, які можуть переходити із одного безмієлінового волокна у сусіднє. У світловому мікроскопі ні мезаксона, ні межі між нейролемоцитами не видно, безмієлінове волокно здається суцільним пучком осьових циліндрів, обгороджених масою цитоплазми, яка міс-тить ядра. Швидкість передачі нервового імпульсу у безмієлінових волокнах менша, ніж у мієлінових. Це пояснюється тим, що у безмієлі-нових волокнах хвиля деполяризацїї рухається по всій плазмолемі, не перериваючись. Відомо, що у внутрішніх органах рух, секреція та інші функції здійснюються повільніше, а тому осьові циліндри безмієліно-вих волокон не ізольовані так чітко один від одного, як це має місце у мієлінових. Розділ 3 Загальна гістологія
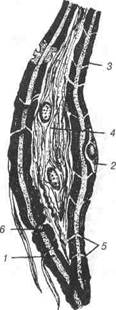
Рис. 74. Схема ультраструктури безмієлінового нервового волокна: 1- ядро; 2-цитоплазма нейролемоцита; 3-4 — аксони нейронів, занурених у цитоплазму нейролемоцита (3-повністю, 4 — частково); 5-мітохондрії; 6-гра-нулярна ендоплазматична сітка; 7-мікротрубочки; 8-колагенові волокна ен-доневрію; 9-базальна мембрана; 10-мезаксон. Мієлінові (від лат. туеііпит — мозок) нервові волокна локалізу-ються як у центральній, так і у периферійній нервовій системі, тобто у складі головного та спинного мозку, а також периферійних нервах. Це товсті волокна діаметром від 3 до 25 мкм, що складаються із осьо-вого циліндра, мієлінової оболонки, нейролеми та базальної мембрани (рис. 75). Осьовий циліндр — це відросток нейрона, яким частіше бу-ває аксон, але може бути і дендрит. Він складається з нейроплазми, яка В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія містить поздовжньо орієнтовані нейрофіламенти і нейротрубки, а та-кож мітохондрії. Осьовий циліндр вкритий аксолемою (продовження плазмолеми нейрона), яка забезпечує проведення нервового імпульсу. В кожному мієліновому волокні лише один осьовий циліндр, навколо якого почергово, ланцюжком розміщуються нейролемоцити.
Рис. 75. Гістоструктура мієлінового нервового волокна: 1-осьовий циліндр-аксон; 2-нейролемоцит; 3-мієлінова оболонка; 4-сполучна тканина; 5-насічки; 6- вузол (перехват Ранв'є). У процесі розвитку мієлінового волокна осьовий циліндр занурю-ється в нейролемоцит, втягуючи його плазмолему і утворюючи глибоку складку (рис. 76). Цю подвійну складку (дуплікатуру) нейролемоцита називають мезаксоном. У процесі подальшого генезу волокна, нейро-лемоцит (швановська клітина) повільно обертається навколо осьового циліндра, в результаті чого мезаксон багато разів огортає його. Цитоплазма нейролемоцита і його ядра залишаються на перифе-рії, утворюючи нейролему волокна. Таким чином, мієлінова оболонка утворюється із щільно й концентрично нашарованих навколо осьово- Розділ 3 Загальна гістологія го циліндра завитків мезаксона, які є пластинками мієлінового шару (рис. 77).
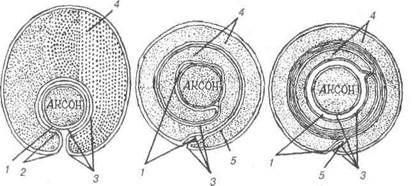
Рис. 76. Схема гістогенезу мієлінового волокна: 1-контакт аксолеми та плазмолеми нейролемоцита; 2-порожнина мезаксона; 3-аксолема та плазмолема нейролемоцита; 4-цитоплазма нейролемоцита; 5-мезаксон.
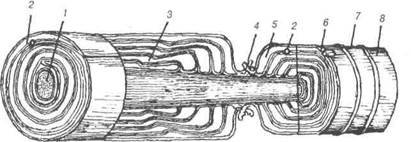
Рис. 77. Схема будови мієлінового волокна: 1-аксон; 2-мезаксон; 3-насічки; 4-вузол (перетяжка Ранв'є); 5-цитоплазма нейролемоцита; 6-ядро нейролемоцита; 7-невролема; 8-ендоневрій. Кожний завиток мезаксона відповідає ліпідним шарам двох лист-ків плазмолеми нейролемоцита. На його середині та поверхні на уль-траструктурному рівні можна виявити темні лінії, утворені білковими молекулами. Насічки мієліну (Шмідта-Лантермана) відповідають тим місцям, де завитки мезаксону розсунуті цитоплазмою нейролемоци- В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологгя, ембріологія та. Насічки можуть мати напрямок як до центру, так і на периферію, у місцях насічок мієлін не переривається. Мієлінова оболонка волок-на складається з багатьох нейролемоцитів, які, в місцях контакту між собою утворюють вузли (перетяжки Ранв'є). Переривність мієлінової оболонки має велике значення для транспортування до осьового ци-ліндра необхідних речовин, сприяє обміну іонами та впливає на швид-кість проведення нервового імпульсу. Ділянка нервового волокна між двома вузлами називається між-вузловим сегментом, який відповідає одній гліальній клітині. На по-вздовжньому розрізі мієлінового волокна поблизу вузла є ділянка, у якій завитки мезаксона послідовно контактують з осьовим циліндром. Місця прикріплення найглибших завитків найбільш віддалені від вуз-лів, а всі наступні — поступово наближаються до них. Це пояснюється тим, що мезаксон нашаровується у процесі росту і осьового циліндра і нейролемоцитів. Таким чином, перші шари мезаксона коротші за останні. Краї двох сусідних лемоцитів, що контактують у ділянці вуз-ла, утворюють інтердигітації, які перекривають ділянку перемички і називаються «вузловою перетяжкою». Міелінові волокна центральної нервової системи у своїй будові мають ряд особливостей. їх оболонку утворюють типові олігоден-дроцити, у них відсутні інтердигітації, насічки Шмідта-Лантермана, відсутня сполучнотканинна базальна мембрана, роль якої виконує нейроглія. Таким чином, у мієліновому волокні лише один осьовий циліндр, мезаксон, закручений щільними шарами, і утворює товстий шар мієлі-ну. Порівняно великий діаметр осьового циліндра, наявність перехва-тів та добре розвинений мієліновий шар забезпечують швидке та точне проведення нервового імпульсу. Синапси Однобічна передача нервового імпульсу в межах рефлекторної дуги зумовлена спеціалізованим контактом двох нейронів, що назива-ють міжнейронним синапсом (від лат зупарзіз - з'єднання). Морфоло-гічно у складі синапсу розрізняють пресинаптичний і постсинаптич-ний полюси, між якими є синаптична щілина. Зустрічаються синапси із хімічною та електричною передачами. Функціонально розрізняють два види синапсів — збудливі та гальмівні. Розділ 3 Загальна гістологія Морфологічна класифікація їх залежить від того, які частини нейро-нів контактують між собою (рис. 78). Аксодендричні -— аксон першого нейрона передає нервовий імпульс на дендрит другого. Аксосоматич-ні — аксон першого нейрона передає імпульс на перикаріон другого. Ак-соаксонні — терміналі аксона першого нейрона закінчуються на аксоні другого. Очевидно, аксоаксонні синапси виконують гальмівну функцію. Крім останніх, найбільш поширених міжнейронних контактів, між де-якими нейронами зустрічаються дендросоматичні та дендродендричні синапси. Таким чином, будь яка частина нейрона може утворювати між-нейронний синапс з будь-якою частиною іншого нейрона.
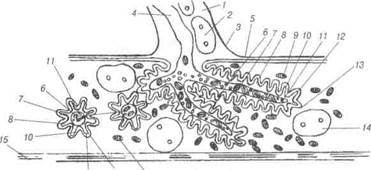
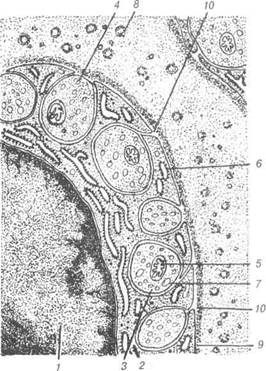
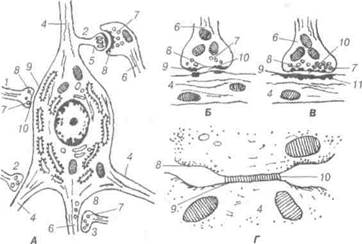
Рис. 78. Схема ультрамікроскопічної будови різних типів синапсів: А —цитотопографіясинапсів;Б —синапсгальмівноготипу;В —сииапсзбудливого типу; Г — електротонічний синапс; 1-аксосоматичний синапс; 2-аксодендричний синапс; 3-аксоаксонний синапс; 4-дендрити; 5-дендритний шипик; 6-аксон; 7-синаптичні пухирці; 8-пресинаптична мембрана; 9-постсинаптична мембрана; 10-синаптична щілина; 11-постсинаптичні структури. Пресинаптичний полюс синапса утворений термінальною гілочкою аксона тієї нервової клітини, яка передає імпульс. Цей полюс, здебіль-шого, розширений у вигляді гудзика, вкритий пресинаптичною мемб- 13-8-305 В.П. Новак, Ю.П. Бичков, М.Ю. Пшипенко Цитологія, гістологія, ембріологія раною. У ньому містяться мітохондрії та синаптичні пухирці, вкриті мембраною і з вмістом певних хімічних речовин, так звані медіатори. Останні сприяють передачі нервового імпульсу на постсинаптичну частину міжнейронного контакту. Синаптичні пухирці бувають різних розмірів від 30 до 150 нм. Це маленькі прозорі та великі електронно-щільні, прозорі, що містять щільну гранулу, а також хімічні речовини, які називаються медіаторами. Участь в передачі нервового імпульсу іншому нейрону відбувається за рахунок транспортування шляхом ек-зоцитозу в синаптичну щілину медіатора. Медіаторами можуть бути ацетілхолін (холінергічні синапси), норадреналін та адреналін (адре-нергічні синапси), а також інші речовини — серотонін, глутамінова кислота, нейротензин, ангіотензин, речовина Р, енкефалін (у збудливих синапсах) дофамін, гліцин, гамма-аміномасляна кислогпа (є медіатора-ми гальмівних синапсів). Пресинаптична мембрана містить електронно-щільні частинки, які разом із нейрофіламентами утворюють пресинаптичну решітку для пухирців. У холінергічних синапсах ці пухирці дрібні і прозорі, а також зустрічаються великі та електронно-щільні, синаптичні пухирці адренергічних синапсів великі, прозорі і морфологічно характеризу-ються наявністю електронно-щільної гранули. Постсинаптична мембрана холінергічних синапсів містить «холін-рецепторний» білок — рецептор медіатора, чим зумовлена дія остан-нього на постсинаптичну мембрану. При взаємодії рецептора медіато-ра з ацетилхоліном утворюються конформаційні зміни його молекул, які призводять до зміни проникності мембрани та генерації нервового імпульсу в нейроні. Медіатори гальмівних синапсів (дофамін, гліцин) не збільшують проникність мембрани для іонів, а зменшують її, стабі-лізуючи мембранний потенціал, а, отже, гальмують генерацію нервово-го імпульсу. Синаптична щілина заповнена тканинною рідиною, в якій міс-тяться електронно-щільні частинки та ниткоподібні структури, що розміщуються на поверхнях обох синаптичних мембран. Очевидно, ці структури містять одночасно пре- і постсинаптичні мембрани. При проходженні нервового імпульсу до закінчення пресинаптичного не-йрона синаптичні пухирці зливаються із пресинаптичною мембраною, їхній вміст виливається в щілину, і медіатор діє на постсинаптичний нейрон. Мембрана пухирців використовується повторно. Розділ 3 Загальиа гістологія Електротонічні синапси утворюються при щільному приляганні нейролем двох нейронів, переважно, їх дендритів і перикаріона. Це, так звані, закриті, електричні синапси (безпухирцеві). Останні не ма-ють синаптичної щілини. У вищих тварин такі синапси зустрічають-ся рідко. 3. 5.5 Нервові закінчення Термінальні апарати нервових волокон — нервові закінчення за їх функціональним значенням поділяють на три види: рецептори, ефек-тори та міжнейронні синапси. Рецептори (чутливі нервові закінчення) — спеціалізовані тер-мінальні структури дендритів нейронів, пристосовані до сприйнят-тя подразнень, що надходять до організму. Залежно від локалізації і специфічності нервової регуляції життєдіяльності організму рецеп-тори поділяють на екстерорецептори (сприйняття подразнень із зовнішнього середовища) та інтерорецептори (подразнення власних тканин організму). Різновидом інтерорецепторів є пропріорецепто-ри — нервові закінчення у м'язах та сухожиллях, які беруть участь у регуляції рухів і положенні тіла у просторі. Залежно від природи подразнень, останні поділяють на термо-, механо-, баро-, хемо-, но-ціорецептори та інші. Чутливі нервові закінчення надзвичайно різноманітні за структур-ною організацією. Вони поділяються на вільні нервові закінчення, які складаються лише із термінальних розгалужень осьового циліндра дендрита чутливого нейрона та невільні — крім осьового циліндра, містять також клітини нейроглїї. Якщо невільні рецептори оточені сполучнотканинною капсулою, їх називають капсульованими; ті не-вільні рецептори, що не мають сполучнотканинної капсули, мають на-зву некапсульовані. Рецепторні закінчення або рецепторно-нейральні синапси складаються із спеціалізованих клітин, здатних реагувати на подразник і викликати деполяризацію плазмолеми зв'язаного з ним дендрита аферентного нейрона. Прикладом вільного рецептора можуть бути деревоподібні та ку-щеподібні нервові закінчення епітеліальної тканини. При формуванні таких закінчень, мієлінові нервові волокна, підходячи до епітеліаль-ного пласта, втрачають мієлінову оболонку, а їхні осьові циліндри утворюють кінцеві розгалуження, що розміщуються між окремими В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія епітеліоцитами. За функцією це, як правило, термо- та ноціорецепто-ри. Вільні нервові закінчення можуть обплітати у вигляді кошика во-лосяні фолікули, реєструючи зміщення у просторі окремих волосин, відіграючи роль механорецепторів. До складу рецепторного апарату багатошарових епітеліїв належать поодинокі чутливі епітеліальні клітини, так звані дотикові епітелгоци-ти Меркеля. Ці електронно-пр.озорі клітини мають сплющені ядра та осміофільні гранули. До їх базальної частини прилягають чутливі не-рвові закінчення у вигляді дисків (дотикові меніски Меркеля). Нейро-епітеліальні клітини Меркеля мають постійне місце в епітеліальному шарі, незважаючи на постійний рух епітеліоцитів від базального до дистального полюса. Клітини Меркеля — типовий приклад нейроепі-телію, епітеліоцити якого пристосувалися до рецепції, вони виконують функцію механорецепції. Надзвичайно різноманітні рецепторні апарати сполучної тка-нини представлені двома групами невільних нервових закінчень. У капсульованих рецепторах нервові терміналі аферентних волокон, як правило, оточені нейролемоцитами і допоміжними елементами спо-лучнотканинного походження. До капсульованих чутливих закінчень, залежно від будови, належать: тільця Фатер-Пачіні, Гольджі-Маццоні, Мейснера, кінцеві колби Краузе (генітальні тгльця). Найбільш поширені рецептори сполучної тканини — це пластин-часті тгльця Фатер-Пачіні (рис. 79). У їх складі розрізняють внутріш-ню колбу та зовнішню капсулу. Навколо розгалужень осьового цилін-дра (термінальний відділ чутливого нервового волокна), яке втратило мієлінову оболонку, скупчуються видозмінені нейролемоцити, що і утворюють внутрішню колбу. Капсула тілець Фатер-Пачіні склада-ється із великої кількості сполучнотканинних пластинок, утворених фібробластами та спірально орієнтованими пучками колагенових во-локон. На межі внутрішньої колби та зовнішньої капсули розміщують-ся клітини, які, очевидно, визначаються як гліальні. Ці гліоцити утво-рюють синапси із розгалуженням осьового циліндра. Є припущення, що нервовий імпульс генерується в умовах зміщення зовнішньої кап-сули щодо внутрішньої колби. Тільця Гольджі-Маццоні менші від тілець Фатер-Пачіні, мають тон-ку капсулу і, порівняно, велику внутрішню колбу Ці структури вико-нують функцію барорецепції у шкірі, серозних та слизових оболонках. Розділ 3 Загальна гістологія Тільця Мейснера (дотикові) — це рецептори тактильної чутливос-ті, які локалізуються у сосочковому шарі дерми. їх олігодендроцити орієнтовані перпендикулярно до поверхні шкіри і мієлінові волокна, втрачаючи мієлінову оболонку контактують із поверхнею гліальних клітин колби. Колагенові волокна сполучнотканинної капсули можуть заповнювати простори між нейролемоцитами і терміналями чутливо-го нервового волокна.
Рис. 79. Схема будови капсульованих рецепторів: А — тільце Фатер-Пачіні; Б — дотикове тільце Мейснера; 1-зовнішня капсу-ла (сполучнотканинні клітини); 2-внутрішня колба (нейрогліальні клітини); 3-осьовий циліндр. Кінцеві колби Краузе є механо- і терморецептпорами кон 'юнкгпиви, язика, а видозмінені структури (генітальні тільця) — зовнішніх стате-вих органів. Характерною особливістю будови колб Краузе є дуже тон-
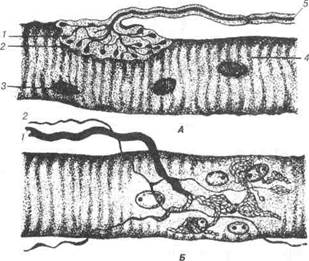
ка сполучнотканинна капсула. Мієлінове волокно, входячи в капсулу, втрачає мієлінову оболонку і закінчується колбоподібним розширен-ням. Генітальні тільця мають подібну будову, за винятком того, що в капсулу проникає не один осьовий циліндр, а декілька, які, розгалужу-ючись, утворюють сіточку безмієлінових волокон. Нервово-сухожильні веретена (сухожильні органи Гольджі) утво-рені товстими мієліновими волокнами, які, підходячи до колагенових волокон, втрачають мієлінову оболонку і утворюють численні розга-луження, обплітають сухожильні пучки. Подібну будову мають тіль-ця Руффіні, що локалізуються у глибоких шарах дерми та підшкірній жировій тканині. Нервово-сухожильні веретена, як і тільця Руффіні вважають механорецепторами, які сприймають взаємне зміщення ко-лагенових волокон. Нервово-м'язові веретена— це чутливі нервові закінчення поперечно-посмугованої м'язової тканини (рис. 80). Вони здатні сприймати зміну довжини м'язового волокна і швидкість цієї зміни. Нервово-м'язові веретена містять кілька інтрафузальних м'язових во-локон, вкритих загальною сполучнотканинною капсулою. До складу веретена входять два товстих по 17 мкм центральних м'язових волокна та чотири тонких по 8 мкм. Екваторіальна частина товстих волокон за-повнена скупченням ядер — «ядерна сумка». Нервово-м'язове веретено має два типи нервових волокон. Одні утворюють спіральні закрутки, що оточують екваторіальну частину товстих інтрафузальних (внутрішньо-веретенних) м'язових волокон — це кільцево-спіральні «анулярні» за-кінчення. Друга група — закінчення чутливих волокон, в своєму складі містить як кільцево-спіральні, так і гроноподібні терміналі, що розміщу-ються по одному з кожного боку кільцеподібних. Перші — реагують на зміну довжини м'язового волокна та її швидкість зміни, другі — лише на зміну довжини м'язового волокна. На обох полюсах м'язових воло-кон локалізуються терміналі рухових нервових волокон. Ефектори — структури, утворені закінченням аксонів нейроци-тів, які бувають рухові та секреторні. Нервово-м'язові закінчення у скелетних м'язах утворені терміналями аксонів нейроцитів, рухових ядер спинного та моторних — головного мозку. При наближенні до м'язового волокна мієлінове волокно втрачає мієлінову оболонку, його осьовий циліндр розгалужується і разом із саркоплазмою міосимплас-та занурюється у м'язове волокно (рис. 81). Роздгл 3 Загальна гістологія
Рис. 80. Схема будови нервово-м'язового веретена: 1-моторні бляшки екстрафузальних м'язових волокон; 2-моторні бляш-ки інтрафузальних м'язових волокон; 3, 4-ядерна сумка (скупчення ядер); 5-м'язовий нерв. У цій ділянці утворюється аксом 'язовий синапс, де аксолема віді-грає роль пресинаптичної мембрани, а сарколема м'язового волокна є постсинаптичною мембраною (рис. 82). Медіатор цих синапсів — аце-тилхолін, який при збудженні вивільняється і через пресинаптичну мембрану проникає у синаптичну гцілину На поверхні постсинаптич-ної мембрани ацетилхолін зв'язується із холінорецепторами і стає хі-мічним сигналом для збудження м'язового волокна. В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
12 13 9 Рис. 81. Схема будови моторної бляшки: 1-цитоплазма нейролемоцита; 2-ядро; 3-нейролема; 4-осьовий циліндр; 5-сар-колема; 6-кінцеві гілочки нервового волокна в повздовжньому та поперечно-му розрізах; 7-мітохондрії аксоплазми; 8-первинний синаптичний простір; 9-саркосоми; 10-вторинний синаптичний простір; 11-синаптичні пухирці; 12-пресинаптична мембрана; 13-постсинаптична мембрана; 14-ядро моторної бляшанки; 15-міофібрила.
Рис. 82. Ефекторне нервове закінчення (моторна бляшка): А — вигляд у профіль: 1,2 — терміналі мієлінового нервового волокна; 3-ядра м'язових волокон; 4-м'язове волокно; 5-мієлінове волокно; Б — вигляд звер-ху: 1- мієлінове волокно; 2- безмієлінове волокно; 3-ультратермінальне во-локно (те, що входить до моторної бляшки і закінчується на іншій). Розділ 3 Загальна гістологія Еферентні закінчення у гладенькій м'язовій тканині мають прості-шу будову. Окремі нервові закінчення на поверхні гладеньких міоци-тів утворюють варикозні терміналі. Медіатор цих закінчень — ацетил-холін, або адреналін.
Розділ 4 Дата добавления: 2015-12-15 | Просмотры: 1239 | Нарушение авторских прав |
 В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія