|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
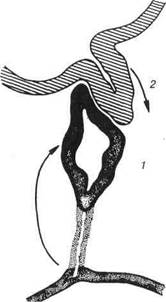
Центральні регуляторні утворення ендокринної системиГіпоталамус (пуроіпаіатуз) — вищий центр ендокринних функ-цій. Він контролює і об'єднує усі вісцеральні функції організму та ен-докринні механізми регуляції з нервовими. Займає базальну частину проміжного мозку і оточує своєю речовиною нижню половину третьо-го мозкового шлуночка. У сірій речовині гіпоталамуса знаходиться 32 пари ядер, розміщених в передній, середній і задній зонах. Упередній зоні знаходяться дві пари ядер, що складаються із вели-ких пептидохолінергічних нейросекреторних клітин. Це супраоптичні та паравентрикулярні ядра. їх клітини мають добре розвинені органе-ли — гранулярну ендоплазматичну сітку, комплекс Гольджі, вони за-безпечують синтез і виділення гормонів. У цитоплазмі нейросекретор-них клітин виявляють специфічні гранули, що містять підготовлені до виведення біологічно активні речовини. Клітини супраоптичних ядер продукують гормон вазопресин, він ви-кликає скорочення міоцитів судин, чим зумовлює підвищення тиску кро-ві; регулює обмін води, оскільки впливає на зворотне всмоктування води Розділ 4 Спеціальна гістологія в сечових канальцях нирок. 3 урахуванням цієї' дії вазопресин називають антидіуретичним гормоном. Також важливу роль відіграє вазопресин у регуляції температури тіла, діяльності серцево-судинної системи. Нейросекреторні клітини паравентрикулярних ядер синтезують окситоцин, який сприяє скороченню міоцитів матки та молочної за-лози. Гормони супраоптичних та паравентрикулярних ядер по аксонах нейросекреторних клітин нагромаджуються у задній частці гіпофіза, потім надходять у кров. Середня зона гіпоталамуса містить дрібні пептидоадренергічні клі-тини, вони об'єднуються в аркуатне, дорсомедіальне, вентромедіальне, супрахиазматичне ядра та преоптичну зону. Клітини цих ядер синте-зують біологічно активні речовини, названі ліберинами і статинами, які впливають на клітини передньої частки гіпофіза. Ліберини і статини об'єднують спільною назвою — релізінг-факторів. Ліберини стимулю-ють продукцію і виведення у кров гормонів гіпофіза до секреції від-повідних тропних гормонів, які впливають на клітини-цілі. Крім того, гіпоталамус надсилає свої імпульси до регульованих ефекторів прямо по симпатичних або парасимпатичних нервах останніх, без опосеред-нення гіпофізом, тобто парагіпофізарно. Нейросекреторна діяльність гіпоталамуса, в свою чергу, зазнає впливу вищих відділів головного мозку, особливо лімбічної системи, мигдалевих ядер, гіпокампа та епіфіза. Гіпофіз (від гр. пурорпузіз сегеБгі- під, рпуотаі — рости) належить до центральних регуляторних органів ендокринної системи. Гіпофіз продукує гормони, дії яких спрямовані на регуляцію функції перифе-рійних залоз ендокринної системи, а також на обмін речовин клітин організму неендокринної природи. Розвиток гіпофіза відбувається за рахунок взаємодії двох ембріо-нальних зачатків — епітеліального та нейрального. Із ектодермального епітелію, що вистеляє ротову ямку зародка, утворюється гіпофізарна кишеня, вона розвивається у напрямку закладки головного мозку і дає початок аденогіпофізу. Назустріч гіпофізарній кишені з боку проміж-ного міхура зародка головного мозку формується виріст, який пере-творюється у лійку третього шлуночка мозку. Нейроглія дистального кінця лійки внаслідок розростання утворює нейрогіпофіз. Прокси-мальна частина лійки звужується і стає гіпофізарною ніжкою, вона зв'язує гіпоталамус з гіпофізом (рис. 117).
Рис. 117. Схема виникнення зародка гіпофіза із двох його головних джерел: 1-випин даху ротової бухти (гіпофізарна кишеня Ратке); 2-нижній відділ тре-тього мозкового шлуночка. Диференціювання епітеліальної гіпофізарної кишені починається розростанням передньої стінки гіпофіза, яка перетворюється в пере-дню частку гіпофіза: задня стінка цієї кишені залишається вузькою се-редньою (проміжною) часткою. Між передньою та середньою частками (за винятком гіпофіза коня) зберігається залишок порожнини гіпофізарної кишені у вигляді вузької гіпофізарної щілини. Таким чином, ембріональний розвиток зумовив будову гіпофіза. Він складається із двох частин: адено- та не-йрогіпофіза. До складу аденогіпофіза входять передня (дистальна), проміжна та туберальна частки. Четвертою часткою гіпофіза (рис. 1 18) є не-йрогіпофіз, який за ноходженням є нейрогліальним. Передня частка утворена розгалуженими епітеліальними тяжа-ми — трабекулами, проміжки між останніми містять пухку сполучну Роздгл 4 Спеціальна гістологія тканину та синусоїдні капіляри. Трабекули утворені залозистими клі- тияами-аденоцитами, серед яких мають місце три різновиди.
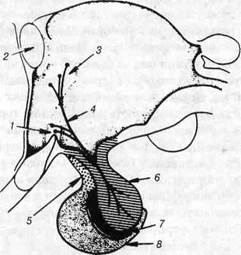
Рис. 118. Схема гіпоталамо-нейрогіпофізарного тракту: 1-супраоптичне ядро; 2-передня комісура; 3-паравентрикулярне ядро; 4-гіпо-таламогіпофізарний тракт; 5-горбкова частина; 6-задня частина; 7-проміжна частка; 8-передня частка. Одні з них знаходяться на периферії трабекул, у цитоплазмі міс-тять секреторні гранули, які інтенсивно сприймають барвники. Ці аде-ноцити називають хромофільними ендокриноцитами. Вони становлять до 40% клітинної маси передньої частки аденогіпофіза. Інпіі клітини займають середню частину трабекули й не мають чітких меж. їх цитоплазма не містить гранул, слабо сприймає барвни-ки, тому такі клітини називають хромофобами. Хромофобних клітин близько 60%. Хромофільні аденоцити здатні сприймати або кислі барвники, або лужні, тому перші відносять до ацидофільних, другі — до базофільних. Найбільший розмір мають базофільні аденоцити, середній — аци-дофільні, найменші — хромофобні клітини. Ацидофільні ендокриноцити округлої або овальної форми, розміщу-ються групами. В їх цитоплазмі знаходяться великі щільні білкові гра- В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія нули, що сприймають кислі барвники — еозин, кислий фуксин, оранж Ж. Центральну частину ацидофільних ендокриноцитів займає ядро, до якого прилягає помірно розвинений комплекс Гольджі, мітохондрії ве-ликого розміру дуже розвинена гранулярна ендоплазматична сітка, що має паралельне розміщення цитомембран. Серед ацидофільних ендокри-ноцитів розрізняють соматотропоцити. Вони продукують соматотроп-ний гормон (СТГ), який впливає на білковий обмін, чим стимулює ріст тіла. У соматотропоцитах секреторні гранули шароподібної форми, а їх діаметр 300-400 нм. Другим різновидом ацидофільних ендокриноцитів є мамотропоцити, вони продукують лактотропний гормон — ЛТГ, або пролактин. Мамотропоцити відрізняються великими цитоплазматич-ними гранулами, розмір яких 400-700 нм. Основне значення ЛТГ (про-лактину) полягає в активізації біосинтезу молока в молочній залозі. Крім того, пролактии продовжує функціонування жовтого тіла в яєч-нику. За кількістю ацидофільні ендокриноцити досягають 30-36% усіх хромофільних аденоцитів передньої частки гіпофіза. Усі ацидофільні ендокриноцити продукують гормони, що за своєю хімічною природою є поліпептидами. Базофільні ендокрітоцити гіпофіза містять гранули, що сприйма-ють основні барвники. Відносна кількість цих клітин становить 4-10% від загальної кількості хромофільних аденоцитів передньої частки. Се-ред базофільних ендокриноцитів розрізняють гонадотропні та тиро-тропні клітини. Гонадотропні становлять 2,2-3,5% від загальної кількості клітин аденогіпофіза. Це великі округлі клітини, більшість їх мають елек-тронно негативне зображення добре розвиненого комплексу Гольджі у вигляді кільцеподібної структури, що має назву макула. Секреторні гранули величиною 200-300 нм, сприймають такі барвники, як альціа-новий синій, альдегід-фуксин та ін. Ендоплазматична сітка цих клітин має вигляд невеликих міхурців або вузьких канальців. Мітохондрії па-личкоподібні, а їх матрикс має підвищену електронну щільність. Гонадотропоцити синтезують фолікулостимулюючий гормон (ФСГ-фолітропін), який впливає на проліферацію сперматогоній сім'яників та фолікулярних клітин яєчника. Другий — лютеїнізуючий гормон (ЛГ-лютропін), його функція полягає у стимуляції жовтого тіла яєчника та стимуляції статевих гормонів самця інтерстиціальни-ми клітинами сім'яника (рис. 119). Розділ 4 Спеціальна гістологія
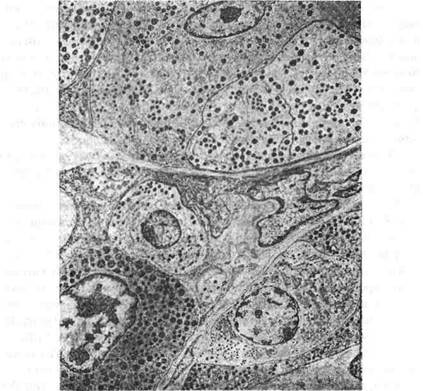
Рис. 119. Електронограма передньої частки гіпофіза х 5450: а-соматотропоцит; б-мамотропоцит; в-кортикотропоцит. Тиротропні клітини характеризуються кутоподібною або непра-вильною формою. Цитоплазма тиротропоцитів містить чітку грану-ляцію діаметром 80-150 нм, яка сприймає альдегід-фуксин, альдегід-тіонин, альціановий синій та інші барвники. Кількість тиротропоци-тів порівняно невелика й становить у пацюків приблизно 1,8-2,9% від загальної кількості секреторних клітин аденогіпофіза. Тиротропоци-ти продукують тиротропний гормон — тиротропін, який впливає на щитоподібну залозу. Крім названих ацидофільних та базофільних ендокриноцитів, передня частка гіпофіза містить третю групу хромофільних клітин, 20 - 8-305 В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія названих кортикотропоцитами. Ці клітини займають переважно центральну зону передньої частки гіпофіза. Вони продукують білко-вий адренокортикотропний гормон — АКТГ-кортикотропін, який під-вищує гормоноутворюючу функцію кіркової частини наднирників. Кортикотропоцити мають неправильну багатокутну форму, їх ядра складаються із окремих часток. Мітохондрії та ендоплазматична сітка добре розвинені. Секреторні гранули мають розмір 100-200 нм і ви-гляд мембранних міхурців з щільною серцевиною. Між мембраною і серцевиною залишається світлий простір. Вважається, що група хромофобних ендокриноцитів є збірною і містить аденоцити з різним ступенем розвитку, диференціювання та різного фізіологічного значення. Серед хромофобних клітин знаходяться, так звані, фолікулярно-зі-рчасті клітини, проте їх природа та значення не з'ясовані. Ці клітини здат-ні формувати мікрофолікулярні структури з відкладенням у них секрету Проміжна частка гіпофіза в препаратах, фарбованих гематоксилі-ном та еозином, має вигляд базофільної смужки епітелію. її клітини здатні продукувати білковий або слизовий секрєт, який нагромаджу-ється, утворюючи фолікулярні кісти. Функціональне значення про-міжної частки гіпофіза достовірно установлено лише для нижчих по-йкілотермних хребетних тварин, у яких меланостимулюючий гормон (МСГ) має провідне значення у регуляції забарвлення тіла. Проміжна частка аденогіпофіза містить меланоцитостимулюючий гормон (мела-ноцитотропін), а також ліпотропін — гормон, що бере участь у метабо-лізмі ліпідів. Є припущення про те, що меланоцитотропін, ліпотропін, а також АКТГ утворюються у головному мозку шляхом розщеплення великої початкової молекули церебрального пептиду на менші фраг-менти, кожний із яких стає одним із названих гормонів. Туберальна частка аденогіпофіза — відділ, що межує з гіпофізар-ною ніжкою та медіальним підвищенням гіпоталамуса. Вона утворена епітеліальними тяжами кубічних клітин із помірно базофільною цито-плазмою. Від останніх, як і від епітелію проміжної частки, у передню частку відходять трабекули; у деяких клітинах туберальних тяжів ви-являють базофільні гранули. Функція клітин туберальної частки гіпо-фіза не визначена. Гіпоталамо-аденогіпофізарне кровопостачання здійснюється пор-тальною (ворітною) судинною системою. Приносні гіпофізарні артерії Розділ 4 Спеціальна гістологія вступають в медіальне підвищення медіального гіпоталамуса і утво-рюють сітку капілярів (первинне капілярне сплетення портальної сис-теми), з якою контактують кінцеві відділи аксонів нейросекреторних клітин аденогіпофізарної зони гіпоталамуса. Капіляри первинного сплетення зливаються у портальні вени, які розміщуються вздовж гіпофізарної ніжки до аденогіпофіза, де розпа-даються на капіляри синусоїдного типу (вторинна капілярна сітка) і розгалужуються між трабекулами паренхіми залози. Синусоїди вторинної капілярної сітки зливаються у виносні вени, в яких кров, збагачена на гормони передньої частки, надходить у за-гальну циркуляцію. Нейрогіпофіз (задня частка) утворена клітинами епендими, які на-зивають пітуїцитами. За формою вони веретеноподібні або зірчасті, мають численні тонкі відростки, що досягають адвентицп кровоносних судин або базальної мебрани капілярів. Нейрогіпофіз акумулює вазопресин та окситоцин, які продуку-ються великими пептидохолінергічними нейросекреторними клітина-ми переднього гіпоталамуса. Аксони цих нейросекрєторних клітин досягають нейрогіпофіза, закінчуються великими терміналями, названі тільцями Херінга, де на-громаджуються секреторні гранули окситоцину та вазопресину. Термі-налі контактують з капілярами, у які й надходять гормони. Епіфіз (від гр. ері - над, рпуотаі — рости) (шишкоподібне тіло) один із центральних органів ендокринної системи. Знаходиться епіфіз між півкулями великого мозку та мозочком. Зовні він обмежений сполучнотканинною капсулою, її прошарки про-никають всередину органа, розгалужуються і поділяють його парен-хіму на часточки. Паренхіма епіфіза містить секретоутворюючі клі-тини — пінеалоцити та гліальні клітини — гліоцити, які відносять до астроцитної глії. Гліоцити виконують опорну функцію, їх відростки мають тісний зв'язок з сполучнотканинною стромою. Пінеалоцити займають центральну частину часток, вони більші від гліоцитів, мають полігональну форму, світле ядро з вєликими ядерця-ми. Від пінеалоцитів відходять розгалужені довгі відростки, які тісно контактують з відростками гліоцитів. Відростки пінеалоцитів біля ка-пілярів утворюють булавоподібні розширення, у складі яких містяться 20* В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія осміофільні гранули. Серед пінеалоцитів виділяються світлі пінеало-цити з бідною на секреторні включення гомогенною цитоплазмою; та темні пінеалоцити меншого розміру з ацидофільними або базофільни-ми гранулами в цитоплазмі. Вважають, що світлі та темні пінеалоцити не самостійні різновиди, а являють собою клітини, що знаходяться у різному функціональному стані. Цитоплазма пінеалоцитів містить численні мітохондрії, розвинений комплекс Гольджі, лізосоми, міхурці агранулярної ендоплазматичної сітки, рибосоми та полісоми. Гліальні клітини, переважно, знаходяться на периферії часток. їх цитоплазма без видимих структур, ядра ущільнені. Під електронним мікроскопом у цитоплазмі виявляють мікрофіламенти діаметром 5-6 нм. Поряд з пінеалоцитами і гліоцитами зустрічаються фіброцити, ма-крофаги, лаброцити, лімфоцити, плазмоцити та пігментні клітини. Функція епіфіза складна й різноманітна. Він затримує розвиток статевої системи, гальмує статеву функцію. Остання зумовлюється тим, що пінеалоцити продукують серотонін, який в епіфізі перетво-рюється у мелатонін. Як вважають, цей нейроамін пригнічує секрецію гонадоліберину гіпоталамусом та гонадотропінів передньої частки гіпофізів. В той же час пінеалоцити продукують ряд білкових гор-монів, зокрема антигонадотропін, який послаблює секрецію лютро-піну передньої частки гіпофіза. Поряд з антигонадотропіном пінеа-лоцитами синтезується значна кількість регуляторних пептидів, яка досягає 40. Епіфіз забезпечує регуляцію фотоперіодичності роботи органів і систем організму, його циркадних ритмів — коливання активності клі-тин у зв'язку із зміною дня та ночі. Механізм реагування епіфіза на зміни освітленості пов'язаний із сприйняттям ним подразнень від сіт-ківки ока по симпатичних нервових стовбурах. Як показують порівняльно-анатомічні дані, епіфіз виникає філо-генетично з тім 'яного ока, яке до цього часу існує в функціональному стані у новозеландської ящірки - гаттерії. Воно затягнене тонкою шкі-рою, містить сітківку, рогівку та кришталик.(рис. 120). Пінеальна залоза виникає у вигляді вип'ячування та виросту до-рсальної стінки третього мозкового шлуночка. Зачаток епіфіза у вели-кої рогатої худоби з'являється вперше у 7-тижневих зародків, а парен-хіма відмежовується після 21-тижневого ембріонального віку. Вікова інволюція епіфіза настає у ранньому віці. Частина пінеа-
Розділ 4 Спеціальна гістологія лоцитів при цьому атрофується, строма розростається, у ній нагрома-джуються вапняні нашарування — мозковий пісок.
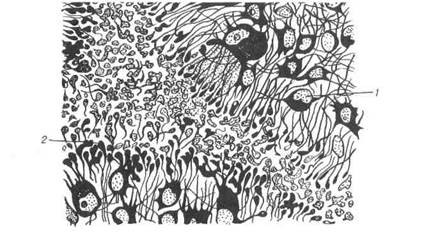
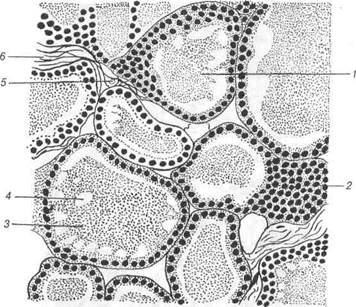
Рис. 120. Епіфіз: 1-пінеалоцит; 2-булавоподібиі розширення відростків пінеалоцитів. 4.5.2 Периферійні ланки ендокринної системи Щитоподібна залоза (від лат. §1апс1иіа іпугеоісіеа). Перенхіма щитоподібної залози містить клітини двох типів: тиро-цитів та паратироцитів. Тироцити розвиваються із вип'ячування ентодермального епітелію вентральної стінки передньої кишки між першою та другою парами зя-брових кишень. Епітелій утворює пухкі сіті трабекул, які галузяться. 3 них формуються замкнені фолікули, а в проміжки між останніми вростає мезенхіма із кровоносними судинами та нервами. Мезенхіма утворює строму Паратироцити - парафолікулярні клітини, розвиваються з нейро-бластів нервового гребеня. Вони залягають поодинці в стінці фолікула між базальною основою тироцитів і базальною мембраною, проте не досягають своєю апікальною поверхнею просвіту фолікула. Крім того, парафолікулярні клітини знаходяться в міжфолікулярній сполучній тканині. їх характеристика буде наведена нижче. В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія Зовні щитоподібна залоза обмежена сполучнотканинною капсу-лою, її прошарки спрямовані всередину й поділяють орган на частки. Структурно-функціональною одиницею щитоподібної залози є фолі-кул. Це мікроскопічний міхурець, стінка якого утворена одним шаром клітин — тироцитів, що лежать на базальній мембрані. Фолікули розділяються прошарками пухкої сполучної тканини, які супроводжуються кровоносними та лімфатичними капілярами, що обплітають фолікули, а також нервові волокна. В сполучнотканинних прошарках знаходяться групи компактних тироїдних епітеліальних клітин. Форма тироцитів тісно пов'язана з функціональною активністю щитоподібної залози. При помірній функціональній активності тиро-цити мають кубічну форму та кулясті ядра. Колоїд, що утворюється тироцитами, у вигляді гомогенної маси заповнює просвіт фолікула. На апікальній поверхні тироцитів містяться мікроворсинки. Бічні поверх-ні сусідніх тироцитів формують десмосомні контакти. Плазмолема ба-зальної поверхні цих клітин утворює інвагінації. Коли функціональна активність тироцитів підвищується, вони набувають призматичної форми, зростає кількість і висота мікро-ворсинок. Інтрафолікулярний колоїд при цьому стає рідким і міс-тить численні резорбційні вакуолі. Одночасно базальна поверхня тироцитів набуває складчатості, що збільшує стикання тироцитів з перифолікулярним простором. При гіпофункції щитоподібної зало-зи висота тироцитів зменшується, вони стають шюскими, а їх ядра набувають витягнутої форми, діаметр фолікулів збільшується, коло-їд стає ущільненим. Цитоплазма тироцитів містить добре розвинені мембранні струк-тури гранулярної ендоплазматичної сітки, комплекс Гольджі, мітохон-дрії, тобто органели, що мають відношення до білкового синтезу. Секреторна функція тироцитів складна й відбувається у дві фази: продукції та виведення гормонів. Фаза продукції полягає у поглинанні через базальну поверхню амінокислот, у тому числі тирозину, йоду та інших мінеральних іонів, деяких вуглеводів, води з крові. У ендоплаз-матичній сітці синтезуються молекули тироглобуліну. Утворені сполу-ки поступово переміщуються у зону комплексу Гольджі, де до поліпеп-тидної основи приєднуються вуглеводні компоненти і утворюються везикули, що містять тироглобулін. Розділ 4 Спеціальна гістологія Цей процес завершується в апікальній частині тироцита, звідки тироглобулін шляхом екзоцитозу надходить всередину фолікула, на-громаджуючись у вигляді колоїду (рис.121,122).
Рис. 121. Щитоподібна залоза коня: 1-фолікул; 2-стінка фолікула; 3-колоїд; 4-вакуоля; 5-капіляр; 6-сполучна тканина. Йод поглинається тироцитами з крові у вигляді іонів, які під вли-вом ферменту пероксидази перетворюються у атомарний. Це відбу-вається на апікальній поверхні тироцита та його мікроворсинок. До тирозинів, що знаходяться у складі тироглобіну, приєднується спо-чатку по одному атому йоду, утворюються монойодтирозини, потім по другому, і монойодтирозини переходять у дийодтирозини, які об'єдну-ються у тетрайодтиронін (тироксин). Поряд з останнім утворюється і трийодтиронін. Фаза виведення. Тироцити своєю апікальною поверхнею шляхом ендоцитозу фагоцитують частини інтрафолікулярного колоїду, які всередині цитоплазми перетворюються у внутрішньоклітинні краплі В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія колоїду. Лізосоми з'єднуються з ними, після їх розщеплення утворю-ються тиреоїдні гормони, які через базальну частину тироцита надхо-дять у загальний кровообіг.
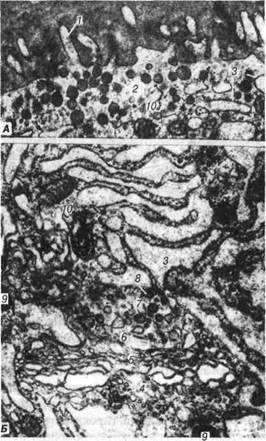
Рис. 122. Електронограма фолікулярної клітини щитоподібної залози: А- апікальна частина клітини: 1-мікроворсинки; 2-апікальні гранули; Б — ор-ганели, які беруть участь у секреції тиреоглобуліну: 3-розтягнуті цистерни гранулярної ендоплазматичної сітки; 4-комплекс Гольджі; 5-транспортні мі-хурці; 6-пресекреторні грануля; 7-секреторні гранули; 8-облямовані міхурці; 9-лізосоми; 10-мітохондрії. Розділ 4 Спеціальна гістологія Паратироцити (кальцитоніноцити - клітини-«К») мають більший розмір ніж тироцити, полігональну або неправильну форму. У цито-плазмі кальцитоніноцитів добре розвинена гранулярна ендоплазма-тична сітка, комплекс Гольджі. На відміну від тироцитів, вони не по-глинають йод. Паратироцити утворюють нейроаміни (норадреналін та серотонін) і синтезують олігопептидні гормони — кальцитонін та соматостатин. Існують два різновиди клітин. У перших переважають дрібні сильно осміофільні гранули. Ці клітини продукують кальцито-нін. Цитоплазма другого типу клітин містить великі, проте слабо осмі-офільні гранули. Ці клітини продукують соматостатин. Щитоподібна залоза іннервується волокнами симпатичної та пара-симпатичної нервової системи. Кровопостачання відбувається за раху-нок гемокапілярів, що відходять від сонної артерії. Прищитовидна залоза — відіграє важливу роль у регулюванні ме-таболізму кальцію. Вона розвивається як потовщення епітелію третьої та четвертої пар зябрових кишень. Зовні паращитовидна залоза вкрита сполучнотканинною капсулою, остання найбільшого розвитку досягає у великої рогатої худоби та коней. Залежно від співвідношення парен-хіми та сполучної тканини, у паращитовидній залозі сільськогосподар-ських тварин виділяють такі архітектонічні типи. 1. Компактний — орган складається їз суцільної маси паренхі-матозних клітин: тонкі сполучнотканинні елементи супроводжують лише кровоносні судини, проте розчленування вони не викликають. Цей тип характерний для собак. 2. Сітчастий тип — сполучнотканинні перекладини та тяжі парен-хіми мають сіткоподібне розміщення: зустрічається у коней та дрібних жуйних. 3. Часточковий — залозисті клітини утворюють округлі часточки, які повністю обмежені сполучною тканиною; він характерний для па-ращитовидних залоз свині. 4. Губчастий тип — характеризується значним розвитком сполуч-нотканинних прошарків; інколи спостерігається у коней та великоїро-гатоїхудоби. Перенхіма прищитовидної залози складається із трабекул-епітеліальних тяжів або скупчень епітеліальних ендокринних клі-тин — паратироцитів, відокремлених тонкими прошарками пухкої сполучної тканини з численними капілярами. В.П. Новак, Ю.П. Бичков, М.Ю. Пклитгенко Цитологія, гістологія, ембріологія Розрізняють головні та оксифільні паратироцити. Головні парати-роцити містять базофільну цитоплазму, периферія її багата на полісо-ми, добре розвинений комплекс Гольджі, мітохондрії. При посиленні секреторної активності прищитовидної залози го-ловні клітини збільшуються у розмірі. Серед головних паратироцитів розрізняють світлі та темні. Цитоплазма світлих паратироцитів міс-тить глікоген. Вважається, що головні, світлі та оксифільні паратиро-цити не є самостійними різновидами паратироцитів, а лише віковим або функціональним станом клітин. Паратироцити продукують білковий гормон — паратирин, який впливає на кісткову тканину — депо нагромадження кальцію в орга-нізмі, стимулює звільнення останнього, що призводить до підвищення рівня цього іона в крові, а в кістках настає часткова демінералізація. Таким чином, паратирин та кальцитонін парафолікулярних клітин щитовидної залози є антагоністами. Прищитовидна залоза за принци-пом зворотного зв'язку реагує на коливання рівню кальцію в крові. Паратироцити мають рецептори, здатні безпосередньо сприймати пря-мий вплив іонів кальцію на них. Іннервуються паращитовидні залози симпатичними та парасимпатичними нервами. Надниркові залози є парними органами. До їх складу входить кір-кова (інтерреналова) тамозкова (супрареналова) частини, об'еднані в один орган. Кіркова частина розвивається з целомічного епітелію. Ці клітини проникають у підлеглу мезенхіму і формують тяжі. їх великі окси-фільні клітини утворюють первинну (фетальну) кору. В подальшому первинна кора обростає дрібними базофільними клітинами целоміч-ного епітелію кореня брижі, з яких утворюється дефінітивна кора. Фе-тальна кора залишається до народження, потім зникає. Мозкова частина розвивається з клітин нервового гребеня, які мі-грують у вентральному напрямку, стають залозистими. Ці клітини спо-ріднені з солями хромової кислоти, тому їх називають хромофінними. Найбільша їх частина проникає у центральну закладку кори і перетво-рюється у мозкову речовину. У зародка хромофінні клітини спочатку містять лише норадреналін, а адреналін з'являється на пізніших стаді-ях ембріогенезу. Зовні надниркові залози обмежені сполучнотканинною капсулою, її зовнішня частина щільна, а внутрішня пухкіша. Від капсули всере- Розділ 4 Спеціальна гістологія дину органа проходять тонкі прошарки, які поряд з капілярами та не-рвами залягають між тяжами паренхіми. Під капсулою знаходиться тонкий прошарок дрібних епітеліальних клітин, за їх рахунок регене-рує кіркова речовина. Кіркова частина утворена поверхневою — дуговою, середньою — пучковою і глибокою — сітчастою зонами. Дугова зона утворена клітинами, переважно, циліндричної форми з овальними або круглими ядрами. Ці ендокриноцити вклинюються між собою і утворюють дугоподібні структури, що характерно для над-ниркових залоз коней, в меншій мірі — для інших тварин. Цитоплаз-ма цих клітин містить агранулярну ендоплазматичну сітку у вигляді дрібних міхурців, між якими знаходяться рибосоми, добре розвинений комплекс Гольджі та мітохондрії. Ендокриноцити дугової зони продуку-ють гормон альдостерон — мінералокортикоїдний гормон, що регулює вміст натрію в організмі, крім того, мінералокортикоїди посилюють за-пальні процеси. Між дуговою та пучковою зонами знаходиться вузька смужка дрібних малодиференційованих клітин. її називають проміжною або суданофільною зоною. Ці клітини вважаються джерелом фізіологічної регенерації пучкової та сітчастої зон. Пучкова зона найбільша. Ендокриноцити, що входять до її складу, мають великі розміри, кубічну або призматичну форму, утворюють тяжі. На поверхні, що межує з капілярами, знаходяться мікроворсинки. Цитоплазма цих клітин багата на краплі ліпідів. Мітохондрії великі, з кристами у вигляді звивистих трубок, які розгалужуються. Добре роз-винена агранулярна ендоплазматична сітка, мають місце рибосоми. Серед ендокриноцитів поряд з світлими мають місце темні, з ущіль-неною цитоплазмою та незначним вмістом ліпідних включень, проте з підвищеним рівнем рибонуклеопротеїдів. Темні клітини містять роз-винену агранулярну та гранулярну ендоплазматичні сітки. Світлі та темні клітини є різновидами функціонального стану ендокриноцитів. Ендокриноцити пучкової зони синтезують глюкокортикоїдні гор-мони (кортизон, ггдрокортизон, кортикостерон), які регулюють обмін вугдеводів, білків, ліпідів, стимулюють енергетичний обмін, пригнічу-ють запальні процеси. Ендокриноцити сітчастої зони кубічної, полігональної або округ-лої форми, утворюють розгалужені пучки. Кількість темних клітин В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія збільшується, а ліпідних речовин зменшується. Клітини сітчастої зони синтезують андрогенстероїдний гормон, подібний до тестостерону, частково — естрогени та прогестерон (рис. 123).
Рис. 123. Надниркова залоза коня: 1-капсула; 2-мозкова речовина; 3-кровоносна судина; 4-нерв; 5-сітчаста зона; 6-пучкова зона; 7-клубочкова зона; 8-кіркова рєчовина. Мозкова речовина міститься у центральній частині наднирника. До її складу входять хромофінні клітини, симпатичні нейрони, не- Роздгл 4 Спеціальна ггстологія рвові волокна та синусоїдні капіляри. Хромофінні клітини великого розміру, округлої або полігональної форми. Серед них розрізняють епінефроцити та норепінефроцити. Епінефроцити — світлі клітини, містять секреторні гранули, продукують адреналін. Цитоплазма норе-пінефроцитів має темний вигляд, містить секреторні гранули гормону норадреналіну. Норадреналін та адреналін відносять до катехоламінів. Адреналін посилює роботу серця, бере участь в регуляції вуглеводного обміну. Норадреналін є медіатором нервового збудження, звужує стін-ку кровоносних судин та підвищує кров'яний тиск, впливає на нейро-секреторну функцію гіпоталамуса. Незначні скупчення клітин мозкової речовини наднирника можуть знаходитися поряд із симпатичними гангліями, тому їх називають па-рагангліями. Васкуляризація мозкової та кіркової речовини наднирника здій-снюється загальним кровопостачанням. Артерії, що надійшли до над-ниркових залоз, розгалужуються на артеріоли, що утворюють густу субкапсулярну сітку; від неї відходять капіляри у кіркову частину. В сітчастій зоні капіляри входять у мозкову речовину, де набувають вигляду синусоїдів і зливаються у венули, які переходять у венозні сплетення мозкової речовини. Поряд з ними у мозкову речовину вхо-дять також артерії. У мозковій частині розгалуження кровоносних су-дин таке, що кожний хромафіноцит одним кінцем контактує з артері-альним капіляром, а другим — з венозним синусоїдом. Кров із кори та мозкової речовини надходить у воротну вену печінки. Іннервуються надниркові залози нервовими імпульсами по черев-них та блукаючих нервах. Дата добавления: 2015-12-15 | Просмотры: 765 | Нарушение авторских прав |