|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Теоретические сведения. Целый ряд грамположительных бактерий образует устойчивые к внешним воздействиям покоящиеся структурыЦелый ряд грамположительных бактерий образует устойчивые к внешним воздействиям покоящиеся структуры, называемые эндоспорами(от лат. spora — семя, посев). Эндоспоры формируются внутри вегетативных клеток бактерий, принадлежащих к родам Bacillus, Sporolactobacillus, Clostridium, Desulfotomaculum, Sporosarcina, Thermoacthomyces. Все эти микроорганизмы образуют толстую клеточную стенку грамположительного типа, что, по-видимому, является необходимым для спорообразования. Эндоспоры чрезвычайно устойчивы к таким факторам, как нагревание, УФ-облучение, действие химических дезинфектантов, растворителей и к высушиванию. В природе образование спор помогает клеткам избегать гибели при истощении субстрата или высушивании, воздействии радиации или химических веществ. Обычно спора в клетке закладывается одна, однако известны случаи формирования до пяти спор в одной бактериальной клетке. Обычно спорообразование начинается, когда бактерии испытывают недостаток питательных веществ или когда в среде в большом количестве накапливаются продукты обмена веществ бактерий. Поэтому споры можно рассматривать как приспособление организма для выживания в неблагоприятных условиях среды. В разных типах материнских клеток (или спорангиях) споры могут залегать по-разному. Тип спорообразования (бациллярный, клостридиальный или плектриднальный) часто помогает в идентификации неизвестной культуры (рис. 20). Иногда образовавшаяся спора столь велика, что расширяет спорангий в середине или с одного конца. В активно спорулирующей культуре почти все клетки образуют споры.
Рис. 20. Типы спорообразования у бактерий. 1 – бациллярный, 2 - клостридиальный, 3 – плектидиальный. Формирование спор зависит от условий роста. Споры могут оставаться живыми в условиях, когда вегетативные клетки, то есть клетки, не образовавшие споры, погибают. Большинство спор хорошо переносит высушивание, многие споры нельзя убить даже кипячением в течение нескольких часов. Для их уничтожения требуется температура пара 120°С при давлении его 1 атм (1,01 -105 Па). При этих условиях споры погибают через 20 мин. В сухом состоянии они погибают лишь при сильном нагревании (до 150—160°С) в течение нескольких часов. Споры отдельных видов бактерий отличаются особенной термоустойчивостью. В процессе образования спор осуществляется синтез особого соединения — дипиколиновой кислоты (пиридин-2,6-дикарбоновая кислота), обычно отсутствующей у вегетативных клеток бактерий. Дипиколиновая кислота может составлять 10—15% массы сухой споры. Показано, что это вещество накапливается в центральной части споры — ее цитоплазме, образуя с ионами кальция комплекс, который вместе с повышенным содержанием других катионов (магния, марганца и калия) обеспечивает пребывание спор в состоянии покоя и их термоустойчивость. Общая схема спорообразования может быть представлена в следующем виде (Рис. 21). В результате неравномерного деления бактериальной клетки, сопровождающегося впячиванием цитоплазматической мембраны, наблюдается обособление части нуклеоида с небольшой частью цитоплазмы. Образовавшаяся проспора затем покрывается цитоплазматической мембраной бактериальной клетки. Таким образом, внутри клетки бактерии возникает новая клетка — проспора, окруженная двумя мембранами. Затем между мембранами образуется кортикальный слой, или кортекс, состоящий из особых молекул пептидогликана. Дальнейшее развитие споры заключается в образовании нескольких слоев споровых покровов и ее созревании. Споровые покровы формируются в основном из вновь синтезированных особых белков, а также липидов и гликолипидов. Электронно-микроскопическое изучение ультратонких срезов спор многих бактерий показало, что поверх покровов споры образуется еще одна структура — экзоспориум, часто состоящий из ряда слоев и имеющий подчас разнообразную «лепную» форму. Диаметр споры приблизительно равен диаметру клетки, в которой она образовалась, или несколько превышает его. У некоторых бактерий спора формируется на конце клетки, которая при этом несколько расширяется, приобретая вид барабанной палочки. У других бактерий спора образуется в центре клетки, и последняя либо не меняет формы (род Bacillus), либо расширяется в середине, принимая вид веретена (род Clostridium). После созревания споры клеточная стенка вегетативной части клетки разрушается, и спора выходит в окружающую среду. При попадании в благоприятные условия спора начинает прорастать. Прорастанию предшествует поглощение спорой воды и последующее набухание. Затем оболочка под влиянием давления, вызванного ростом, разрывается, возникает ростовая трубка. В дальнейшем происходит удлинение освободившегося бактериального организма и, наконец, деление уже удлиненной клетки.
Рис. 21. Формирование эндоспоры спорообразующими бактериями: I — вегетативная клетка; II — инвагинация ЦПМ; III — образование споровой перегородки (септы); IV— формирование двойной мембранной системы образующейся проспоры; V— сформированная проспора; VI — формирование кортек-са; VII — формирование покровов споры; VIII — лизис материнской клетки; IX — свободная зрелая спора; X — прорастание споры; 1— нуклеоид; 2 — цитоплазма; 3 — ЦПМ; 4 — клеточная стенка; 5 — споровая перегородка; 6 — наружная мембрана споры; 7— внутренняя мембрана споры; 8— кортекс; 9— покровы споры. Споры бактерий могут длительное время (десятки, сотни и даже тысячи лет) существовать в покоящемся состоянии. Некоторые эндоспоры остаются жизнеспособными в течение 800 лет (например, споры бацилл сибирской язвы в скотомогильниках), споры актиномицетов — до 7 500 лет. но совершенно уникальным является случай проращивания спор Bacillus cereus, обнаруженных в кишечнике пчелы, найденной в кусочке янтаря, насчитывающего 25 — 30 млн лет. Имеются микроорганизмы, образующие относительно устойчивые к неблагоприятным условиям среды (температура, кислотность, аэрация и др.) покоящиеся клетки — цисты, не являющиеся спорами (Рис 22). Например, азотобактер образует цисты, устойчивые к высушиванию и теплу. Известны и другие группы покоящихся клеток (миксоспоры миксобактерий, акинеты цианобактерий, эндоспоры актиномицетов и др.).
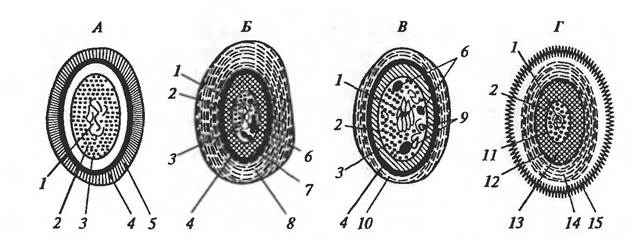
Рис. 22. Строение покоящихся форм прокариот: А — миксоспоры миксобактерий; Б — цисты азотобактера; В - акинеты цианобактерий; Г — эндоспоры; 1 — нуклеоид; 2 — цитоплазма; 3 — ЦПМ; 4 — клеточная стенка; 5 — капсула; 6 — гранулы запасных веществ; 7 — внутренние покровы (интина); 8 — внешние покровы (экзина); 9 — тилакоиды; 10 — чехол; 11 — внутренняя мембрана споры; 12 — наружная мембрана споры; 13 — кортекс; 14 — покровы споры, состоящие из нескольких слоев; 15 — экзоспориум Большинство дифференцированных специализированных структур относится к категории покоящихся форм, назначение которых — обеспечить переживание вида в течение длительного времени в неблагоприятных условиях. Это эндоспоры ряда грамположительных бактерий, цисты азотобактера и миксобактерий, акинеты цианобактерий, экзоспоры отдельных представителей метилотрофных и фототрофных бактерий, экзо- и эндоспоры актиномицетов. После попадания в подходящие условия покоящиеся формы прорастают, давая начало вегетативным клеткам. Некоторые прокариоты, наряду с эукариотичеекими организмами типа простейших, образуют другие виды покоящихся форм, называемые цистами. Циста — это потерявшая подвижность клетка с утолщенной, ослизненной оболочкой, устойчивая к высушиванию и перепадам температур. Цисты характеризуются состоянием покоя со значительно сниженной метаболической активностью. Цисты выполняют три основные функции: 1) защищают популяцию от вредных воздействий окружающей клетку среды, таких, как истощение источника питания, высушивание, неблагоприятный рН или низкое парциальное давление кислорода; 2) служат способом сохранения ДНК; 3) являются способом передачи инфекционного начала от хозяина к хозяину (у патогенов). Обычно прорастание цист стимулируется благоприятными внешними условиями для развития клеток. Цисты встречаются у разных групп эубактерий: азотобактера, спирохет, миксобактерий, риккетсий. У большинства миксобактерий образование цист, называемых также миксоспорами,— закономерная стадия их жизненного цикла (рис. 23, А). После окончания стадии активного размножения клетки миксобактерий собираются вместе и образуют так называемые плодовые тела, представляющие собой массу слизи, в которую погружены клетки, или весьма дифференцированные структуры, поднимающиеся над поверхностью субстрата на простых или разветвленных стебельках (рис. 23, Б). Внутри плодовых тел клетки переходят в покоящееся состояние.
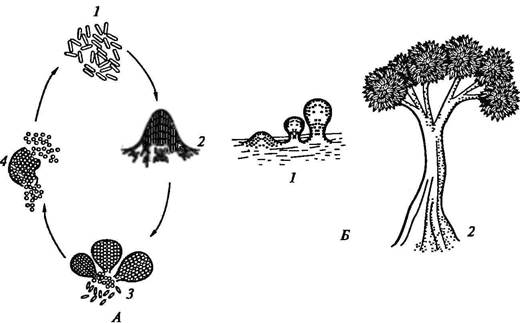
Рис. 23. Цикл развития и плодовые тела некоторых миксобактерий: А — цикл развития Myxococcus: 1 — активно размножающиеся вегетативные клетки; 2 — скопление клеток, предшествующее образованию плодового тела; 3 — плодовое тело; 4 — миксоспоры; Б — плодовые тела: 1 — Myxococcus; 2 — Chondromyces У одних видов цисты могут морфологически не отличаться от вегетативных клеток, у других их образование сопровождается заметными морфологическими и структурными изменениями: происходит утолщение стенки вегетативной клетки, в результате чего формируются оптически плотные, более сильно преломляющие свет, окруженные капсулой укороченные палочки или сферические формы. Образование миксоспор сопровождается синтезом белка. ДНК не синтезируется, а переходит из исходных вегетативных клеток. Генетический аппарат миксоспор может быть представлен тремя или четырьмя копиями хромосомы вегетативной клетки. Цисты миксобактерий более устойчивы к нагреванию, высушиванию, различным физическим воздействиям, чем вегетативные клетки. Покоящимися клетками некоторых цианобактерий, обладающими повышенной устойчивостью к ряду неблагоприятных факторов (высушиванию, пониженным температурам), являются акинеты. Они, как правило, заметно крупнее вегетативных клеток, имеют продолговатую или сферическую форму, гранулированное содержимое и толстую оболочку. Образование акинет происходит в период замедления роста и начинается с увеличения клеточных размеров, при этом в цитоплазме происходит накопление гранул запасных веществ (гликогеновых, полифосфатных и особенно крупных цианофициновых), а также карбоксисом. Одновременно происходит утолщение пептидогликанового слоя клеточной стенки и уплотнение слизистого чехла за счет отложения в нем электронно-плотного фибриллярного материала полисахаридной природы. Оболочки акинет содержат больше липидов и полисахаридов, а цитоплазма — меньше воды, чем вегетативные клетки. В цитоплазме при формировании акинет отмечается увеличение содержания ДНК, рибосом, но уменьшение количества хлорофилла и фикобилиновых пигментов. Тилакоиды образуют сложную сетчатую структуру. Скорость фотосинтеза в акинетах ниже, а дыхание выше, чем в вегетативных клетках. Прорастание акинет происходит иногда вскоре после их образования или только после перенесения в свежую питательную среду и может осуществляться двумя путями: иногда в акинете на одном из полюсов формируется пора, через которую выходит проросток, или же прорастание происходит в результате разрыва оболочки акинеты. У актиномицетов споры являются покоящимися клетками и одновременно репродуктивными структурами. По типу образования они делятся на две группы — эндогенные и экзогенные. Эндогенное образование спор внутри цитоплазмы материнской гифы, обнаруженное у представителей родов Thermoactinomyces и Actinobifida, протекает аналогично описанному выше. У большинства актиномицетов споры формируются экзогенно путем деления гифы перегородками на участки, каждый из которых представляет собой будущую спору. Экзоспоры большинства актиномицетов не содержат каких-либо дополнительных внутренних структур помимо тех, которые наблюдаются в вегетативной клетке. Стенка споры обычно значительно толще, чем стенка гифы, и в ней можно различить несколько слоев разной электронной плотности. Часто клеточная стенка окружена дополнительными наружными покровами. Дата добавления: 2015-12-16 | Просмотры: 1341 | Нарушение авторских прав |