|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
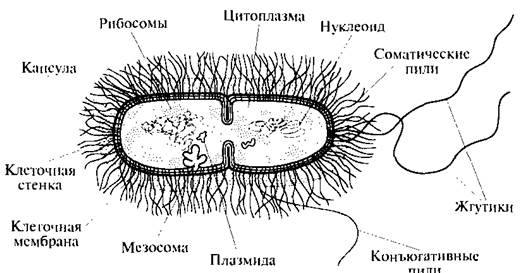
Теоретические сведения. Бактериальные хромосомы были открыты много позднее, чем эукариотические, так как они не обладают свойством конденсироваться в метафазеБактериальные хромосомы были открыты много позднее, чем эукариотические, так как они не обладают свойством конденсироваться в метафазе, что делает хромосомы эукарий столь видимыми. В 40-х годах XX в. были получены первые доказательства спонтанного мутагенеза у бактерий, и это послужило основанием для предположения о наличии у прокариот мутирующих генов — функциональных элементов хромосомы. Почти в то же время А. Эвери с коллегами раскрыл химическую природу генетического материала в экспериментах по трансформации авирулентного штамма пневмококка препаратом ДНК, выделенным из вирулентного штамма, с получением вирулентных клеток. Но в то время эти данные не были восприняты безоговорочно. Потребовались годы до открытия в 1952 г. того факта, что ДНК, а не белок входит в клетку бактерии при фаговой инфекции (А.Д.Херши и М.Чейз), и до расшифровки структуры ДНК в 1953 г. (Дж.Уотсон и Ф. Крик), чтобы факт передачи наследственной информации через ДНК был наконец признан. В 1956 г. внутри бактериальных клеток была обнаружена «ядерная зона», или н уклеоид, где размешена бактериальная хромосома. В 70-х годах стало возможным выделение компактной формы тотальной ДНК из клеток бактерий, что положило начало интенсивным биохимическим исследованиям нуклеиновых кислот прокариот. Состояние суперскрученности бактериальной ДНК было обнаружено в середине 60-х годов, в течение 10 лет после этого были обнаружены ферменты, которые отвечают за сверхспирализацию и раскручивание ДНК (топоизомеразы, гиразы). В целом, по представлениям молекулярной биологии, хромосома содержит: большие молекулы ДНК как носители генетической информации; молекулы РНК, копирующие и передающие информацию с определенных генов; белки, которые репарируют повреждения ДНК, удваивают ДНК и контролируют модели экспрессии генов. Белки также скручивают и складывают ДНК внутри клетки. Бактериальная ДНК обнаружена и кольцевой, и линейной форм. Индивидуальная клетка прокариот может содержать несколько идентичных копий одной хромосомы. У некоторых видов в клетке обнаружены две и даже три неидентичные хромосомы. Когда клетка содержит одну очень большую плазмиду, трудно определить разницу между плазмидой и хромосомой, так как обе молекулы могут нести гены, необходимые для роста (Рис 24).
Рис. 24. Схема делящейся бактериальной клетки Большинство бактерий несет все гены в одной группе сцепления, т.е. на одной хромосоме. Однако появляются свидетельства того, что разные гены могут располагаться и на разных хромосомах. От идеи о том, что прокариоты имеют лишь одну кольцевую хромосому, уже отказались. Более того, число видов с более чем одной группой сцепления генов (одной хромосомой) может оказаться гораздо больше, чем мы представляем на сегодняшний момент, поскольку значительное количество существующих в мире микроорганизмов не выделено и не изучено. Наличие множественных копий некоторых частей хромосомы, а также всей хромосомы (полиплоидия) хорошо известно у эукариот. В дополнение к этому, клетки эукарий могут содержать тысячи копий митохондриальных и хлоропластных геномов, мультикопийные гены и хромосомы находят также и у бактерий. Возможно, клетки используют ДНК как запасное вещество, подобно крахмалу, жиру или гликогену для других организмов. В связи с вышеизложенным, бактериальные и эукариотические хромосомы нельзя больше рассматривать как совершенно разнородные в отношении формы (оба типа могут быть линейными), уровня плоидности (оба типа могут быть полиплоидными) и количества групп сцепления (бактерии, которые часто имеют одну, могут иметь несколько групп; эукариоты, которые обычно имеют несколько, могут иметь только одну группу, как в случае Myrmecia pilosula). Остается самое большое различие между эу- и прокариотами в организации наследственного аппарата — окруженное ядерной мембраной оформленное ядро у эукарий, наличие гистонов и упаковка ДНК в нуклеосомы. У бактерий не найдены пока гистоны и нуклеосомы, поэтому упаковка ДНК у них происходит иначе. С другой стороны, некоторые археи имеют гистоноподобные белки и стабильные нуклеосомы, в то время как некоторые одноклеточные эукариоты лишены и того и другого. Таким образом, на уровне организации хромосом различия между про- и эукариотами могут быть не столь явными, как это ранее предполагалось. При электронно-микроскопическом наблюдении видно, что нуклеоид прокариот, несмотря на отсутствие ядерной мембраны, довольно четко отграничен от цитоплазмы, занимает в ней, как правило, центральную область и заполнен нитями ДНК диаметром около 2 нм. Не исключено, что на выявляемую в электронном микроскопе организацию прокариотной хромосомы большое влияние оказывают условия фиксации препарата. По имеющимся наблюдениям, в живой клетке нуклеоид занимает больше места в цитоплазме. Вся генетическая информация прокариот содержится в одной молекуле ДНК, имеющей форму ковалентно замкнутого кольца. Длина молекулы в развернутом виде может составлять более 1 мм, т.е. почти в 1000 раз превышать длину бактериальной клетки. Длительное время считали, что в распределении нитей ДНК бактериальной хромосомы не прослеживается никакой закономерности. Однако если исходить из того, что молекула ДНК образует беспорядочный клубок, трудно объяснить процесс репликации и последующее распределение образовавшихся хромосом по дочерним клеткам. Специальные исследования показали, что хромосомы прокариот представляют собой высокоупорядоченную структуру, имеющую константу седиментации 1300—2000 S для свободной и 3200—70005 для связанной с мембраной формы. В том и другом случае часть ДНК в этой структуре представлена системой из 20— 100 независимо суперспирализованных петель. В обеспечении суперспирализованной организации хромосом участвуют молекулы РНК. В прокариотной клетке ДНК может находиться и вне бактериальной хромосомы — в плазмидах, но последние не являются обязательными клеточными компонентами. Деление молекулы ДНК (репликация) происходит по полуконсервативному механизму и в норме всегда предшествует делению клетки. С помощью электронного микроскопа установлено, что репликация ДНК начинается в точке прикрепления кольцевой хромосомы к ЦПМ, где локализован ферментативный аппарат, ответственный за репликацию. Часто можно обнаружить, что контакт ДНК с ЦПМ осуществляется посредством мезосом. Репликация, начавшаяся в точке прикрепления, идет затем в двух противоположных направлениях, образуя характерные для кольцевой хромосомы промежуточные структуры. Возникающие дочерние хромосомы остаются прикрепленными к мембране. Репликация молекул ДНК происходит параллельно с синтезом мембраны в области контакта ДНК с ЦПМ. Это приводит к разделению (сегрегации) дочерних молекул ДНК и оформлению обособленных хромосом (рис. 25).
Рис. 25. Механизм распределения бактериальных хромосом: А — бактериальная клетка содержит частично ре пли циро ванную хромосому, прикрепленную к мембране в точке (или точках) репликации; Б — репликация хромосомы завершена. В бактериальной клетке две дочерние хромосомы, каждая из которых прикреплена к ЦПМ. Показан синтез клеточной стенки и ЦПМ; В — продолжающийся синтез мембраны и клеточной стенки приводит к разделению дочерних хромосом. Показано начало деления клетки путем образования поперечной перегородки; J — ДНК; 2 — прикрепление хромосомы к ЦПМ; 3 — ЦПМ; 4 — клеточная стенка; 5 — синтезированный участок ЦПМ; 6 — новый материал клеточной стенки Согласно существующим представлениям суперспирализованные петли соответствуют неактивным в данное время участкам ДНК и находятся в центре нуклеоида. По его периферии располагаются деспирализованные участки, на которых происходит синтез информационной РНК (иРНК), при этом, поскольку у бактерий процессы транскрипции и трансляции идут одновременно, одна и та же молекула иРНК может быть одновременно связана с ДНК и рибосомами. Дата добавления: 2015-12-16 | Просмотры: 912 | Нарушение авторских прав |