|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Динамичность геномаМетоды новой генетики расширили наши знания о структуре генетического материала. Представление о хромосоме как о нитке с бусинами-генами соответствует реальным фактам теперь еще в меньшей степени, чем раньше. Неясно даже, что, собственно, мы должны называть «геном» (см. ниже). Очень важно также, что по современным данным генетический материал намного менее статичен, чем представлялось раньше. И хотя эти новые данные трудно пока как-либо использовать в биологии и патологии человека, мы считаем нужным сделать несколько кратких замечаний относительно динамичности генома. Мобильные элементы и транспозоны. Известно, что початки кукурузы могут иметь мозаичную окраску (рис. 2.93). Генетика этого явления была изучена Барбарой МакКлинток [433]. Она пришла к выводу, что в геноме кукурузы существуют некие «контролирующие элементы», которые могут перемещаться с одного гена на другой, увеличивая их нестабильность. Мозаичная окраска початков у кукурузы обусловлена соматическими мутациями, связанными с присутствием контролирующих элементов. Их характерные особенности были проанализированы в большой серии изящных экспериментов. В течение длительного времени контролирующие элементы у кукурузы казались уникальным исключением. Так продолжалось до 1963 г., когда Тэйлор [524] описал «индуцированные фагом мутации у Е. coli». Этот фаг теперь называется Ми (от 2. Хромосомы человека 141
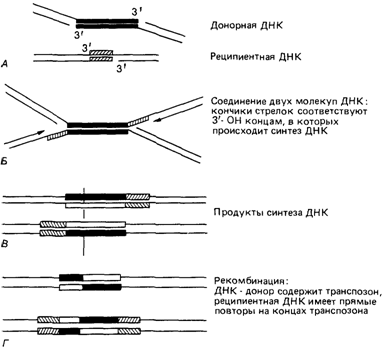
англ. mutator – мутатор). Вскоре после этого Старлингер и Сэдлер [520] описали ISэлементы у бактерий. Мобильные элементы у бактерий [520; 358]. Эти элементы теперь определяют как специфические последовательности ДНК, которые могут неоднократно внедряться в разные сайты генома. У прокариот различают три класса таких элементов: 1) Is-элементы или простые инсерционные последовательности не содержат никаких генов, кроме тех, которые связаны с инсерционной функцией. Длина Is-элементов обычно меньше 2 т. п. н. 2) Тп-элементы (транспозоны). Помимо генов, участвующих в транспозиции, они несут дополнительные гены, например устойчивости к антибиотику. Длина транспозонов обычно больше 2 т. п. н. 3) Эписомы. Они представляют собой сложные самореплицирующиеся структуры, часто содержащие Is- и Тп-элементы. С помощью секвенирования ДНК и других методов было показано, что мобильные элементы обладают следующими свойствами: Is-последовательности несут на концах полные или почти полные инвертированные повторы длиной в 20-40 п. н.; большинство Tn-элементов оканчивается длинными (800-1 500 п. н.) ls-подобными последовательностями. Когда элементы встраиваются в хозяйский геном, они фланкируются короткими повторами (4-12 п. н.) ДНК хозяина (рис. 2.94). Мобильные элементы могут встраиваться во многие сайты генома хозяина с помощью механизма негомологичной рекомбинации, но иногда внедрение бывает строго специфичным. Показано, что при внедрении мобильного элемента встраивается не он сам, а его копия, в то время как исходный элемент остается на своем месте (рис. 2.95). Когда мобильный элемент встраивается в структурный ген, целостность последнего нарушается, что приводит к генной мутации. Кроме того, инсерция мобильного элемента может привести к возникновению хромосомных аберраций, таких, как делеции, дупликации, инверсии и транслокации. Мобильные элементы у эукариот. Как уже указывалось, мобильные элементы впервые были обнаружены у кукурузы. Впоследствии их нашли и у других эукариот, например у дрозофилы [365]. У этого организма инсерционные мутанты возникают с высокой частотой и несут вставки в определенных специфических локусах. Существует три источника мобильных элементов у дрозофилы: а) они могут происходить из генных последовательностей, рассеянных по всему геному. Показано, что некоторые элементы из этих генных последовательностей действительно способны перемещаться; б) вторым возможным источником мобильной ДНК могут быть повторяющиеся последовательности конститутивного гетерохроматина прицентромерных районов хромосом; в) в качестве источника мобильных элементов могут выступать и РНК-вирусы, паразитирующие на дрозофиле. В этом случае с помощью обратной транскриптазы (разд. 2.3.2.2) вирусная РНК транскрибируется в ДНК, а последняя способна встраиваться в геном. Мобильные элементы дрозофилы наряду с индивидуальными особенностями в структурной организации, такими, как наличие инвертированных повторов на концах, обладают общими свойствами с транспозонами бактерий. Они способны с высокой частотой индуцировать генные мутации, причем мутации эти нестабильны и часто ревертируют; их репликация независима; частота возникновения хромосомных аберраций у линий, несущих мобильные элементы, повышена. Описаны также мобильные элементы у дрожжей [358]. Значение мобильных элементов в эволюции. Высказываются предположения, что перенос генов мобильными элементами может служить одним из факторов эволюции. Если помимо классических путей передачи наследственной информации от родителей к потомкам существует еще горизонтальный перенос (даже между отдаленными видами), разнообразие генетических изменений должно резко возрасти. Заметим, что, например, перенос генов от одной бактерии к другой с помощью фага (трансдукция) известен давно, а теперь используется и в 142 2. Хромосомы человека
2. Хромосомы человека 143
генетической инженерии эукариот (включая клетки млекопитающих). Возможно, что такие процессы могут происходить и в природе. Более того, последовательности ДНК, гомологичные глобиновому гену человека, были обнаружены у бобовых растений [312]. Функция такого гена у растений может состоять в том, чтобы «обеспечить кислородом клубеньковые бактерии в ткани». Наличие этого гена может быть объяснено переносом его от насекомых или млекопитающих. Существуют ли мобильные элементы в геноме человека? До сих пор подобные элементы в геноме человека еще не выявлены. Однако, также как и у дрозофилы, у человека имеются рассеянные по геному повторяющиеся фрагменты ДНК (разд. 2.3.1.1), иногда содержащие даже палиндромные последовательности, которые по аналогии могли бы рассматриваться в качестве мобильных элементов. Например, онкогены имеют структурную гомологию с клеточными РНК-вирусами (ретровирусами, разд. 5.1.6); сходные с ретровирусами повторяющиеся элементы идентифицированы в ДНК человека [429]; показано, что вирусная ДНК мутагенна для клеток млекопитающих [1463]. В геноме человека обнаружена особая группа диспергированных повторяющихся последовательностей, так называемые Alu-последовательности. Уже указывалось, что ядерная ДНК человека организована по типу ДНК Xenopus, т.е. состоит из уникальных последовательностей длиной 1-2 т. п. н., перемежающихся повторяющимися последовательностями длиной 0,1-0,3 т. п.н. Мы говорили также, что некоторые из этих последовательностей представляют собой палиндромы, т. е. состоят из комплементарных инвертированных повторов (разд. 2.3.1.1). Однако если в геноме Xenopus эти повторяющиеся последовательности формируют много разных семейств, то у млекопитающих, таких, как грызуны или приматы, они обнаруживают сильную гомологию [505]. У человека ≈ 3-6% всей геномной ДНК приходится на повторяющиеся последовательности длиной 300 п. н. и ≈ 60% таких повторов, как показано рестрикционным анализом, оказываются гомологичными. Общее число копий Alu-последовательностей оценивается в настоящее время в 500 000 на гаплоидный геном, т. е. в среднем одна такая последовательность приходится на каждые 5000 пар оснований, но распределены они неравномерно. Другими словами, примерно одна Alu-последовательность встречается через каждые 2,5 рассеянных по геному ДНК-повторов различного типа, включая инвертированные повторы и палиндромы (рис. 2.96). С обеих сторон они фланкированы обычно короткими прямыми повторами длиной от 7 до 20 пар оснований. В отличие от собственно Alu-последовательностей эти повторы уникальны для разных Alu-последовательностей. Как указывалось выше, такие повторы фланкируют бактериальные транспозоны, как и мобильные элементы эукариот. Именно поэтому был сделан вывод, что Alu-последовательности имеют такое же происхождение, что и мобильные элементы, а фланкирующие их повторы возникли в результате дупликации коротких последовательностей сайта транспозиции. Последовательности типа Alu, как правило, находят в первичных транскриптах РНК, они удаляются в ходе процессинга РНК (разд. 2.3.3.6). Вероятно, что рассеянными по геному эти последовательности оказываются благодаря тому, что их относительно короткие РНК-транскрипты транскрибируются обратной транскриптазой в ДНК, которая встраивается затем в различные участки генома. Поскольку эти последовательности сохранились в ходе эволюции млекопитающих (о чем свидетельствует частичная гомология между приматами, включая человека, и грызунами), они должны иметь важные функции. По аналогии с подобными элементами у других эукариот, таких, как кукуруза и дрозофила, они могут участвовать в регуляции экспрессии генов или рекомбинационном процессе как в зародышевых, так и в соматических клетках. Первым этапом в транспозиции определенной последовательности ДНК должно быть образование ее экстрахромосомных, кольцевых копий. Подобные элементы действительно обнаружены в стареющих фибробластах in vitro [494; 400]. 144 2. Хромосомы человека
Конверсия генов. Еще один относящийся к обсуждаемому предмету феномен давно известен в экспериментальной генетике под названием генной конверсии [122]. Различные данные, полученные при изучении глобиновых генов, позволяют предполагать наличие такого феномена и в геноме человека (разд. 4.3; см. также рис. 2.97). Генная конверсия есть не что иное, как модификация одного из двух аллелей другим, в результате чего гетерозигота Аа, например, становится гомозиготой АА. Винклер, который впервые обсуждал этот феномен более 50 лет тому назад, допускал «физиологическое взаимодействие» аллелей. Однако работы на дрожжах показали, что он связан с атипичной рекомбинацией. Данный процесс иллюстрирован на рис. 2.97. Кроссинговер всегда приводит к разрыву последовательности ДНК в сайте перекреста. Обычно разрыв репарируется, для чего последовательность сестринской хроматиды используется как матрица. Таким образом восстанавливается исходная двойная спираль. Однако иногда репарация осуществляется на матрице гомологичной хромосомы. В этом случае наблюдаются отклонения от обычной сегрегации. Генная конверсия имеет место и в соматических тканях, особенно у растений. Возможно, что в этом случае рекомбинационный процесс протекает атипично. Наличие генной конверсии не является неожиданным, поскольку спаривание гомологичных хромосом в соматических клетках и соматический кроссинговер характерны для многих видов (см., например, [368]). Подобная ситуация может иметь место в некоторых случаях ретинобластомы (злокачественная опухоль глаза у детей) (разд. 5.1.6). Существует ряд доказательств, что генная конверсия происходит в области HLA-тютаусоъ и в кластере глобиновых генов. В этом последнем случае она может быть причиной таких нарушений, как Э-талассемия, HbS или НЬЕ (разд. 4.3). Изменяется ли геном? Насколько стабильны генетическая информация и ее передача? Изучая наследование моногенных заболеваний или таких полиморфных систем, как группа крови АВО или MN, мы не можем не поразиться точности передачи генетической информации, указывающей на стабильность генома. В конце концов напрашивается вывод, что встречающиеся иногда исключения вполне можно объяснить не биологическими факторами, а скорее такими, как, например, ложное отцовство. Единственное, что в какой-то степени ослабляет нашу веру во всеохватывающую надежность наследственных механизмов,-это новые мутации (разд. 5.1), но их частота обычно очень низка, и, кроме того, однажды возникнув, они подчиняются правилам генетической передачи. Однако открытия в молекулярной биологии поставили перед нами важные вопросы. Мы знаем теперь, что гены могут фрагментироваться, перемещаться по геному, конвертировать свои аллели. Они могут встраиваться в наш геном не тем приятным 2. Хромосомы человека 145 способом, который оправдан временем и который практиковали еще наши предки, а например с «бродячим» вирусом. Должны ли мы в связи с этим забыть элементарную генетику и подвергнуть сомнению все ее правила? К счастью, классические правила остаются справедливыми. Арбер в своей Нобелевской лекции, посвященной генетическому обмену, сказал: «Несмотря на то что существует множество естественных механизмов, способствующих обмену между геномами неродственного происхождения, и Е. coli, и высшие организмы преуспели в достижении относительно высокой общей стабильности их генетического аппарата». Новые данные углубляют наше понимание структурной организации генетического материала и механизмов его работы. Несомненно, что они могут помочь в предотвращении наследственных болезней. Но верно и то, что старые правила остаются справедливыми.
146 2. Хромосомы человека Дата добавления: 2015-12-16 | Просмотры: 736 | Нарушение авторских прав |