|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
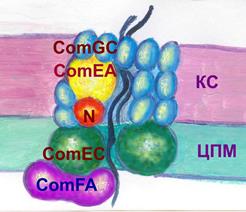
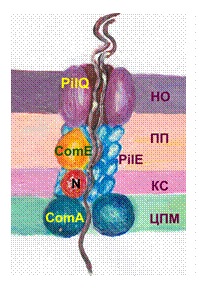
Трансформация. Трансформация - разновидность рекомбинативной изменчивости микроорганизмов, сопровождающаяся переносом ДНК от донора к реципиентучерез окружающую средуТрансформация - разновидность рекомбинативной изменчивости микроорганизмов, сопровождающаяся переносом ДНК от донора к реципиентучерез окружающую среду. В процессе трансформации ДНК-фрагмент погибшей и разрушенной бактерии встраивается в ДНК реципиента, находящегося в состоянии компетенции. Впервые продемонстрирована в 1928 г. Фредериком Гриффитом. Описана как у грамположительных, так и грамотрицательных бактерий родов Bacillus, Haemophilus, Neisseria, Escherichia, Pneumococcus. Для осуществления трансформации необходимы следующие условия: l. Наличие небольшого (до 20 генов) фрагмента двухцепочечной ДНК донорской клетки. Одноцепочечные или меньшего размера ДНК быстро разрушается нуклеазами окружающей среды. 2. Наличие реципиента, находящегося в состоянии компетенции. В естественных условиях захват ДНК клеткой-реципиентом осуществляется в фазу логарифмического роста, когда продуцируется специфические ДНК-связывающие белки, кодируемые com-генами. Состояние проницаемости бактериальных мембран для ДНК называют компетенцией. В лабораторных условиях компетенцию достигают путем обработки химическими веществами, например, добавлением CaCl2 в культуру. Стадии трансформации. На первой стадии трансформации происходит захват ДНК клеткой-рецепиентом. У грамотрицательных микроорганизмов (например, N. gonorrhoeae)ДНК присоединяется к поверхности бактерии и поступает в ее цитоплазму по каналу, пронизывающему наружную мембрану, пептидогликан, периплазматическое пространство и цитоплазматическую мембрану. В области наружной мембраны канал состоит из белка-пиллина PilQ, в пептидогликане и периплазматическом пространстве из Pil E и ДНК-связывающего белка Com E, в цитоплазматической мембране - из белка Сom A. Перед поступлением в канал, состоящий из белка Сom A, одна из двух нитей ДНК разрушается нуклеазой. У грамположительных микроорганизмов ( например, B. subtilis) нет наружной мембраны, и потому транспорт происходит только через клеточную стенку и цитоплазматическую мембрану. В клеточной стенке для прохождения ДНК формируется канал из белка Сom GC и ДНК-связывающего протеина Com EA. На уровне цитоплазматической мембраны одна из нитей ДНК разрушается нуклеазой, а вторая
Рис. 4. Каналы для поступления ДНК в процессе трансформации НО – наружная мембрана, ПП – периплазматическое пространство, КС – клеточная стенка, ЦПМ – цитоплазматическая мембрана
На второй стадии трансформации происходит реципрокный обмен между донорской и реципиентной ДНК, называемый гомологичной рекомбинацией. Рекомбинация требует сходства между генетической информации донора и реципиента и наличия генов бактериальной рекомбинации- recA, B и C. В редких случаях возможна рекомбинация между отдаленными видами. Значение. В естественных условиях трансформация приводит к усилению вирулентности. Используется в микробиологической науке для конструирования генетически модифицированных микроорганизмов, изучения функций генов.
Трансдукция Трансдукция -разновидность рекомбинативной изменчивости микроорганизмов, сопровождающаяся переносом генетической информации от донора к реципиенту с помощью бактериофага. Перенос участков бактериальной хромосомы фагами был открыт в 1951г. Ледербергом и Циндером у Salmonella typhimurium, впоследствииописана у многих родов бактерий: Salmonella, Escherichia, Shigella, Bacillus, Pseudomonas, Vibrio, Streptococcus, Slaphylococcus, Corynebacterium. Капсидная оболочка бактериофага защищает ДНК от действия нуклеаз, поэтому трансдукция, в отличие от трансформации, не чувствительна к нуклеазам. Трансдукцию осуществляют умеренные фаги. Они переносят лишь небольшой фрагмент генома клетки хозяина, и как правило, среди особей одного вида, но возможен и межвидовой перенос генетической информации, если бактериофаг имеет широкий спектр хозяев. В зависимости от исхода взаимодействия фага с бактерией выделяют литические и умеренные фаги. Литические (вирулентные) фаги впрыскиваютнуклеиновую кислоту в клетку и репродуцируются в ней, после чего покидают клетку путем лизиса. Лизогенные, или умеренные фаги, инъецировав свою ДНК в клетку, могут вести двояко: 1) начать цикл репродукции и покинуть клетку путем лизиса; 2) интегрировать свою генетическую информацию в геном бактерии и в его составе передаваться дочерним клеткам. Фаги, встроенные в геном бактерий, называют профагами, а бактерии со встроенными в геном фагами, - лизогенными. В результате действия факторов, прерывающих лизогению (УФ, ионизирующей радиация, химические мутагены), вновь синтезируются вирусные частицы, которые покидают клетку. Примером умеренного фаг является фаг l, поражающий E. coli. Этапы его трансдукции:
a. Неспецифическую (общую). Бактериофаг может встраиваться в любом месте генома бактерии и потому способен переносить любой фрагмент ДНК хозяина. b. Специфическую. Бактериофаг встраивается в строго определенные места генома бактерии, а потому переносит лишь строго определенные фрагменты ДНК. c. Абортивную. Участок бактериальной хромосомы донора, перенесенный бактериофагом, не вступает в рекомбинацию с хромосомой реципиента, а остается вне хромосомы. Происходит транскрипция перенесенной ДНК (на это указывает синтез соответствующего генного продукта), но не репликация. В процессе деления клетки донорский фрагмент переходит только в одну из дочерних клеток и со временем утрачивается.
Конъюгация Конъюгация – разновидность рекомбинативной изменчивости микроорганизмов, сопровождающаяся переносом генетической информации от донора к реципиенту через непосредственный контакт (конъюгационный мостик). В естественных условиях происходит с частой 1:106. Ее существование доказано в 1946 г. Ледербергом и Татумом. Для грамотрицательных микроорганизмов конъюгация - основной механизм горизонтальной передачи генов. Может осуществляться среди особей одного вида, так и разных видов, и способствует передаче таких признаков, как множественная антибиотикорезистентность. Грамположительные микроорганизмы также могут путем конъюгации передавать признаки, однако, основной механизм у них – трансдукция. У грамположительных микроорганизмов клетки-доноры продуцируют адгезивный материал, который способствует агрегации с реципиентом и передаче генетического материала. Для осуществления конъюгации необходимо выполнение следующих условий: 1. Наличие донора. Донор отличается от реципиента присутствием полового фактора F (от fertility - плодовитость), представляющего автономно реплицирующуюся плазмиду (кольцевая двухцепочечная молекула ДНК с массой 45 х 106 Да), содержащую около25 генов, которые детерминируют: 1) процесс независимого от генома удвоения (репликации); 2) особые структуры клеточной поверхности - половые пили, или F-пили, которыеслужат для взаимного узнавания при контакте донора и реципиента и образуют конъюгационный мостик. 2. Наличие реципиента. Не имеет плазмиды F. Исход конъюгации и частота передачи хромосомных признаков зависят от физиологическое состояние F фактора. Физиологические состояния F плазмиды: 1. Автономное. F фактор находится в цитоплазме в свободном состоянии, не интегрирован в бактериальную хромосому и не несет в своем составе хромосомные гены. 2. Интегрированное, или Hfr. F фактор может интегрироваться в определенных местах в бактериальную хромосому, и в таком состоянии носит название эписомы. Донорские клетки, с интегрированным фактором F, обеспечивают высокую частоту переноса хромосомной ДНК, и получили название клеток Hfr (от англ. high frequency of recombinants). 3. Автономное F¢. Интегрированная F плазмида может покидать бактериальную хромосому, захватывая близлежащие гены, таким образом превращаясь в F¢ фактор.
Рис. 5. Физиологические состояния F плазмиды Дата добавления: 2015-12-16 | Просмотры: 788 | Нарушение авторских прав |