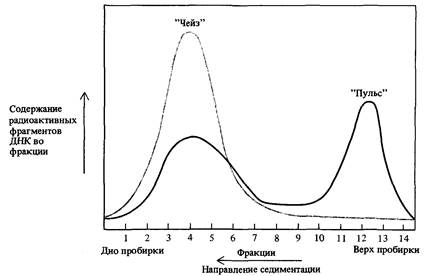
Рис. 13.2. Эксперимент, подтверждающий модель, согласно которой часть новообразованной ДНК Е. coli состоит из небольших одноцепочечных фрагментов, которые в дальнейшем включаются в состав протяженных цепей. Черная кривая отражает распределение по размерам одноцепочечных фрагментов ДНК, наблюдаемое при пульсовом введении метки и последующем центрифугировании в сахарозном градиенте плотности при pH 12 для предотвращения образования водородных связей между комплементарными цепя-
ми. Небольшие фрагменты, содержащиеся в пике, расположенном в верхней части центрифужной пробирки, через некоторое время после удаления из среды радиоактивных предшественников (период «чейз») оказываются включенными в состав протяженных одноцепочечных участков (цветная линия). Разделение основано на том, что большие по размеру одноцепочечные цепи характеризуются более высокой скоростью седиментации, чем небольшие фрагменты.
ДНК. Эту РНК-полимеразу, отличающуюся от ферментов, которые непосредственно участвуют в процессе транскрипции, называют праймазой.
РНК-затравочные участки не сохраняются в структуре зрелой ДНК. После реализации своей функции для инициации репликации ДНК они удаляются за счет проявления 5' ® 3'-экзонуклеазной активности ДНКполимеразы (рис. 13.1, Б и 13.3). После удаления РНК-затравки и ее замещения на фрагмент ДНК, инициация синтеза которого происходит на следующей РНК-затравке, расположенной ближе к области репликативной вилки, между двумя соседними синтезированными фрагментами ДНК остается разрыв. Этот разрыв (отсутствие ковалентной связи между 3'-ОН- и 5'-РО4-концами фрагментов цепи) устраняется при участии фермента - ДНК-лигазы, - направляющего образование фосфодиэфирной связи (рис. 13.4).
Синтез ведущей и отстающей цепей происходит по мере продвижения репликативной вилки вдоль двойной спирали родительской ДНК (см. рис. 13.1). Процесс расплетания двойной спирали и экспонирования