|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Общая схема репликации вирусов (цикл одиночного развития фага, биохимия вирусной инфекции).
Репродукцией (лат. productio - производство) вирусов называют процесс размножения вирусных частиц в чувствительных к ним клетках. В цикле репродукции тех и других вирусов различают четыре стадии: 1) подготовительную, или инициальную, включающую фазы адсорбции вируса на клетке, проникновения и раздевания в клетке; 2) собственно репродуктивную стадию образования структурных белков и вирионных нуклеиновых кислот; 3) сборку вирионов; 4) заключительную, сопровождающуюся выходом зрелых вирусных частиц из клетки. Подготовительная стадия репродукции. Взаимодействие вирусов с клетками определяется наличием у них биологического сродства (тропизма), и прежде всего специфических рецепторов. С них и начинается «узнавание» вирусом чувствительных клеток. Находятся они на поверхности наружной мембраны клеток в большом количестве, не менее 104 - 105. По химической структуре рецепторы разнородны - молекулы-белки, углеводные или липидные компоненты протеидов. Адсорбция этих и других вирусов на соответствующих рецепторах осуществляется с помощью имеющихся у них прикрепительных белков. У простых вирусов прикрепительные белки содержатся в капсидах, у сложных - в суперкапсидах, например в шипах или булавовидных утолщениях у коронавирусов. Следующая за адсорбцией фаза проникновения вируса в цитоплазму клетки реализуется двумя способами - пассивным путем виропексиса или активным путем слияния (интеграции) вирусной оболочки с клеточной мембраной. Виропексис представляет собой своеобразную форму эндоцитоза, при котором вирус проникает в клетку путем впячивания мембраны с образованием вокруг него вакуоли и поэтапным ее слиянием вначале с более крупной цитоплазматической вакуолью, а далее - с лизосомой или же с внутриклеточными мембранами, включая ядерную. Второй, более редкий путь проникновения вирусов в клетку происходит посредством интеграции вирусных оболочек с наружной мембраной. Отмечается он лишь у тех видов вирусов, которые наделены белками слияния. Как и прикрепительные, белки слияния содержатся в капсидах простых и суперкапсидах сложных вирусов. Способность к репродукции вирусы приобретают только после освобождения их нуклеиновых кислот от оболочек, что принято называть фазой раздевания (депротеинизации). Осуществляется раздевание вирусов ферментами поверхностных плазматических мембран клеток-хозяев при слиянии с ними вирусных оболочек, а при виропексисе - ферментами лизосом и внутриклеточных цитоплазматическйх и ядерных мембран. Конечными продуктами раздевания у ряда вирусов являются, правда, не голые нуклеиновые кислоты, а НК, связанные с внутренним вирусным белком (пикорна-, аденовирусы), нуклеокапсидом (вирус гриппа) или сердцевиной (аденовирусы). Собственно репродуктивная стадия. Истинно репродуктивная стадия, включающая фазы транскрипции, трансляции и репликации, у разных групп вирусов и семейств неодинакова. Так, транскрипция, или переписывание нуклеиновых кислот вирусов на иРНК, как первый ее этап, у ДНК-содержащих семейств папова-, адено- и герпесвирусов, репродукция которых происходит в ядре, осуществляется клеточной РНК-полимеразой, а у репродуцирующихся в цитоплазме иридо- и поксвирусов — вирусоспецифической, попадающей в клетку вместе с их геномом. Второй этап реализации генетической информации вирусов - трансляция, в процессе которой синтезируется их белок, начинается с узнавания клеточными рибосомами вирусных иРНК, чему способствуют особые вирусоспецифические инициаторные факторы. В процессе трансляции у вирусов, кодирующих синтез одной длинной иРНК, синтезируется гигантский полипептид-предшественник, впоследствии нарезающийся на несколько различных белков, а у вирусов, кодирующих короткие иРНК, - соответствующее им число полностью созревших белков. Репликация ДНК-содержащих вирусов, или копирование их генома, представленного линейной двухцепочечной структурой, сходна с репликацией ДНК клеток и осуществляется их ДНК-полимеразами. Другими словами, синтез гомологичных нуклеиновых кислот происходит на обеих расплетенных цепях, в результате чего каждый вновь образующийся вирион получает ДНК, состоящую из старой цепи и ее новой копии. Следует, правда, подчеркнуть, что раскручиванию кольцевых двухцепочечных ДНК предшествует разрезание одной из ее нитей, а репликация однонитчатых ДНК-содержащих парвовирусов происходит после синтеза второй цепи ДНК и образования промежуточных двухцепочечных его форм. Репликация вирусных РНК тоже происходит не на родительских, а на промежуточных комплементарных нитях. При этом синтез тех и других нитей РНК, так же как иРНК, осуществляют не клеточные, а вирусоспецифические РНК-полимеразы. Процесс репликации комплементарных нитей РНК у минус-нитевых вирусов хотя и очень схож с процессом синтеза иРНК при транскрипции, но они не аналогичны друг другу. В отличие от относительно коротких иРНК, комплементарных отдельным участкам генома, комплементарно-репликативные РНК считываются с полного генома и по существу представляют собой антигеном. Следовательно, в инфицированных вирусами клетках существует механизм переключения частичного считывания генома (транскрипции) на сквозное (репликацию). Вновь синтезированная при сквозном считывания антигенома геномная РНК может служить матрицей для синтеза новых ее копий или иРНК, т. е. войти в состав вириона или функционировать как иРНК. Сборка вирионов. Сборка вирусных компонентов в корпускулярную частицу - сложный и до конца не выясненный процесс. Хорошо известно лишь то, что при формировании простых вирусов в ее основе лежит самопроизвольный механизм белок-нуклеиновой сборки. У сложно устроенных вирусов сборка происходит поэтапно и во многом зависит от того, где происходит их репликация - в цитоплазме или ядре. Начинается она всегда с формирования нуклеокапсидов (сердцевин) и синтеза суперкапсидных белков. При этом образование первых происходит в специальных структурах клеток, или «фабриках», индуцированных вирусами, а вторых — в полирибосомах, связанных с внутриклеточными мембранами. Затем нуклеокапсиды, глико- и липопротеиды суперкапсидов РНК-вирусов мигрируют в места их сборки, которыми обычно являются мембраны эндоплазматических сетей, а чаще - поверхностные мембраны клеток. Связывание нуклеокапсидов с суперкапсидными субъединицами может происходить без или с участием матриксных мембранных М-белков вирионов, способных к белок-белковым и белоклипидным взаимодействиям. Процесс сборки ДНК-содержащих вирусов еще сложнее. Начинаясь с формирования сердцевин или нуклеокапсидов, протекает через образование неполных форм вирионов с разным содержанием ДНК в капсидах и незрелых вирионов с полностью ненарезанными в них предшественниками полипептидов, свойственных зрелым формам вирионов. При окончательном формировании зрелых вирионов в их внешние оболочки часто включаются липиды, углеводы, белки и даже ферментные системы клеток-хозяев. Заключительная стадия репродукции. Конечным этапом репродукции вирусов является выход сформированных вирионов из инфицированных клеток. При этом возможны два пути их освобождения: 1) отпочковывание от клеток, присущее сложным вирусам, имеющим липопротеидную оболочку; 2) разрушение клеток, которое в основном вызывают безоболочечные простые пикорна-, рео-, парво-, папова- и аденовирусы.
31. Стадии репликации вирусов: адсорбция (рецепторы вирусов), проникновение, депротеинизация вирусной частицы, синтез предшественников вирусных нуклеиновых кислот и белков, сборка вирионов, выход вирусных частиц из клетки.
Репродукцией (лат. productio - производство) вирусов называют процесс размножения вирусных частиц в чувствительных к ним клетках. В цикле репродукции тех и других вирусов различают четыре стадии: 1) подготовительную, или инициальную, включающую фазы адсорбции вируса на клетке, проникновения и раздевания в клетке; 2) собственно репродуктивную стадию образования структурных белков и вирионных нуклеиновых кислот; 3) сборку вирионов; 4) заключительную, сопровождающуюся выходом зрелых вирусных частиц из клетки. Подготовительная стадия репродукции. Взаимодействие вирусов с клетками определяется наличием у них биологического сродства (тропизма), и прежде всего специфических рецепторов. С них и начинается «узнавание» вирусом чувствительных клеток. Находятся они на поверхности наружной мембраны клеток в большом количестве, не менее 104 - 105. По химической структуре рецепторы разнородны - молекулы-белки, углеводные или липидные компоненты протеидов. Адсорбция этих и других вирусов на соответствующих рецепторах осуществляется с помощью имеющихся у них прикрепительных белков. У простых вирусов прикрепительные белки содержатся в капсидах, у сложных - в суперкапсидах, например в шипах или булавовидных утолщениях у коронавирусов. Следующая за адсорбцией фаза проникновения вируса в цитоплазму клетки реализуется двумя способами - пассивным путем виропексиса или активным путем слияния (интеграции) вирусной оболочки с клеточной мембраной. Виропексис представляет собой своеобразную форму эндоцитоза, при котором вирус проникает в клетку путем впячивания мембраны с образованием вокруг него вакуоли и поэтапным ее слиянием вначале с более крупной цитоплазматической вакуолью, а далее - с лизосомой или же с внутриклеточными мембранами, включая ядерную. Второй, более редкий путь проникновения вирусов в клетку происходит посредством интеграции вирусных оболочек с наружной мембраной. Отмечается он лишь у тех видов вирусов, которые наделены белками слияния. Как и прикрепительные, белки слияния содержатся в капсидах простых и суперкапсидах сложных вирусов. Способность к репродукции вирусы приобретают только после освобождения их нуклеиновых кислот от оболочек, что принято называть фазой раздевания (депротеинизации). Осуществляется раздевание вирусов ферментами поверхностных плазматических мембран клеток-хозяев при слиянии с ними вирусных оболочек, а при виропексисе - ферментами лизосом и внутриклеточных цитоплазматическйх и ядерных мембран. Конечными продуктами раздевания у ряда вирусов являются, правда, не голые нуклеиновые кислоты, а НК, связанные с внутренним вирусным белком (пикорна-, аденовирусы), нуклеокапсидом (вирус гриппа) или сердцевиной (аденовирусы). Собственно репродуктивная стадия. Истинно репродуктивная стадия, включающая фазы транскрипции, трансляции и репликации, у разных групп вирусов и семейств неодинакова. Так, транскрипция, или переписывание нуклеиновых кислот вирусов на иРНК, как первый ее этап, у ДНК-содержащих семейств папова-, адено- и герпесвирусов, репродукция которых происходит в ядре, осуществляется клеточной РНК-полимеразой, а у репродуцирующихся в цитоплазме иридо- и поксвирусов — вирусоспецифической, попадающей в клетку вместе с их геномом. Второй этап реализации генетической информации вирусов - трансляция, в процессе которой синтезируется их белок, начинается с узнавания клеточными рибосомами вирусных иРНК, чему способствуют особые вирусоспецифические инициаторные факторы. В процессе трансляции у вирусов, кодирующих синтез одной длинной иРНК, синтезируется гигантский полипептид-предшественник, впоследствии нарезающийся на несколько различных белков, а у вирусов, кодирующих короткие иРНК, - соответствующее им число полностью созревших белков. Репликация ДНК-содержащих вирусов, или копирование их генома, представленного линейной двухцепочечной структурой, сходна с репликацией ДНК клеток и осуществляется их ДНК-полимеразами. Другими словами, синтез гомологичных нуклеиновых кислот происходит на обеих расплетенных цепях, в результате чего каждый вновь образующийся вирион получает ДНК, состоящую из старой цепи и ее новой копии. Следует, правда, подчеркнуть, что раскручиванию кольцевых двухцепочечных ДНК предшествует разрезание одной из ее нитей, а репликация однонитчатых ДНК-содержащих парвовирусов происходит после синтеза второй цепи ДНК и образования промежуточных двухцепочечных его форм. Репликация вирусных РНК тоже происходит не на родительских, а на промежуточных комплементарных нитях. При этом синтез тех и других нитей РНК, так же как иРНК, осуществляют не клеточные, а вирусоспецифические РНК-полимеразы. Процесс репликации комплементарных нитей РНК у минус-нитевых вирусов хотя и очень схож с процессом синтеза иРНК при транскрипции, но они не аналогичны друг другу. В отличие от относительно коротких иРНК, комплементарных отдельным участкам генома, комплементарно-репликативные РНК считываются с полного генома и по существу представляют собой антигеном. Следовательно, в инфицированных вирусами клетках существует механизм переключения частичного считывания генома (транскрипции) на сквозное (репликацию). Вновь синтезированная при сквозном считывания антигенома геномная РНК может служить матрицей для синтеза новых ее копий или иРНК, т. е. войти в состав вириона или функционировать как иРНК. Сборка вирионов. Сборка вирусных компонентов в корпускулярную частицу - сложный и до конца не выясненный процесс. Хорошо известно лишь то, что при формировании простых вирусов в ее основе лежит самопроизвольный механизм белок-нуклеиновой сборки. У сложно устроенных вирусов сборка происходит поэтапно и во многом зависит от того, где происходит их репликация - в цитоплазме или ядре. Начинается она всегда с формирования нуклеокапсидов (сердцевин) и синтеза суперкапсидных белков. При этом образование первых происходит в специальных структурах клеток, или «фабриках», индуцированных вирусами, а вторых — в полирибосомах, связанных с внутриклеточными мембранами. Затем нуклеокапсиды, глико- и липопротеиды суперкапсидов РНК-вирусов мигрируют в места их сборки, которыми обычно являются мембраны эндоплазматических сетей, а чаще - поверхностные мембраны клеток. Связывание нуклеокапсидов с суперкапсидными субъединицами может происходить без или с участием матриксных мембранных М-белков вирионов, способных к белок-белковым и белоклипидным взаимодействиям. Процесс сборки ДНК-содержащих вирусов еще сложнее. Начинаясь с формирования сердцевин или нуклеокапсидов, протекает через образование неполных форм вирионов с разным содержанием ДНК в капсидах и незрелых вирионов с полностью ненарезанными в них предшественниками полипептидов, свойственных зрелым формам вирионов. При окончательном формировании зрелых вирионов в их внешние оболочки часто включаются липиды, углеводы, белки и даже ферментные системы клеток-хозяев. Заключительная стадия репродукции. Конечным этапом репродукции вирусов является выход сформированных вирионов из инфицированных клеток. При этом возможны два пути их освобождения: 1) отпочковывание от клеток, присущее сложным вирусам, имеющим липопротеидную оболочку; 2) разрушение клеток, которое в основном вызывают безоболочечные простые пикорна-, рео-, парво-, папова- и аденовирусы.
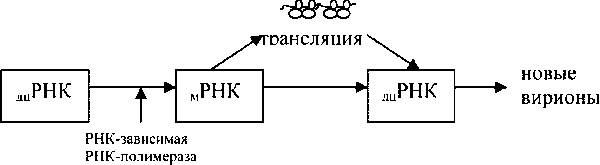
32. Основные типы репликации вирусных геномов: двухцепочечные ДНК -геномы, одноцепочечные (+)ДНК -геномы
Вирусы с двуцепочечной ДНК. Здесь, например, возбудители герпеса и оспы. В зараженной клетке фермент ДНК-зависимая РНК-полимераза считывает (транскрибирует) с генома этих вирусов молекулы мРНК (то есть (+)РНК), которые делают свою обычную работу - направляют синтез белков. Размножением вирусного ДНК-генома занимается фермент ДНК-зависимая ДНК-полимераза. Поскольку геном клетки также представлен молекулами двуцепочечных ДНК, то в ней еще до заражения имеются как ДНК-зависимая РНК-полимераза, так и ДНК-зависимая ДНК-полимераза. В некоторых случаях производством вирусных мРНК и ДНК занимаются клеточные ферменты. Другие же вирусы (более сложные) это крайне ответственное дело никому не передоверяют и используют собственные ферменты. Бывает и так, что транскрипция и репликация вирусного генома - прерогатива "смешанных предприятий", эксплуатирующих как вирусных, так и клеточных "работников". Заканчивается инфекционный цикл, как обычно, "одеванием" генома в защитную одежду и выходом вирионов "в свет". 1-Цепоч ДНК: Попав в клетку, вирусный геном сначала превращается в двуцепочную форму, это превращение обеспечивает клеточная ДНК-зависимая ДНК-полимераза. Далее смотри предыдущий вариант.
33. Основные типы репликации вирусных геномов: (+)-РНК -геномы, двухцепочечные РНК-геномы,
Дата добавления: 2015-12-16 | Просмотры: 1441 | Нарушение авторских прав |