|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
ОРІЄНТОВНА КАРТКА ДЛЯ РОБОТИ З ЛІТЕРАТУРОЮ.
МАТЕРІАЛИ ДЛЯ САМОКОНТРОЛЮ Дайте відповіді на запитання: 1. Яку будову має функціональна одиниця м’язового волокна – саркомер? 2. Які типи скоротливих протофібрил Вам відомі? 3. Який білок, подібний до гемоглобіну, міститься у м'язах? 4. Чому м'язи, у яких переважають червоні волокна, здатні до більш тривалої безперервної активності? 5. Які ознаки властиві тетанічному типу скорочення м'язів? 6. Яку будову має міон?
Тема: «Механізми скорочення та розслаблення скелетних м'язів»
АКТУАЛЬНІСТЬ ТЕМИ Сучасні знання про механізм скорочення м'язового волокна базуються на уявленні про філаменти двох типів, що зсуваються одні відносно інших. Ці уявлення є основою моделі ковзних ниток, запропонованої Г. Хакслі зі співпрацівниками на базі електронно-мікроскопічних досліджень та рентгеноструктурного аналізу. Щоб з'ясувати механізм взаємодії актинових і міозинових філаментів, слід розглянути їхню молекулярну будову. ФОРМУВАННЯ НАВЧАЛЬНИХ ЦІЛЕЙ ЗНАТИ принципи ініціації м’язового скорочення, механізм розслаблення м’яза. ВМІТИ пояснювати принцип дії «шарнірного механізму» ковзання протофібрил м’язового волокна.
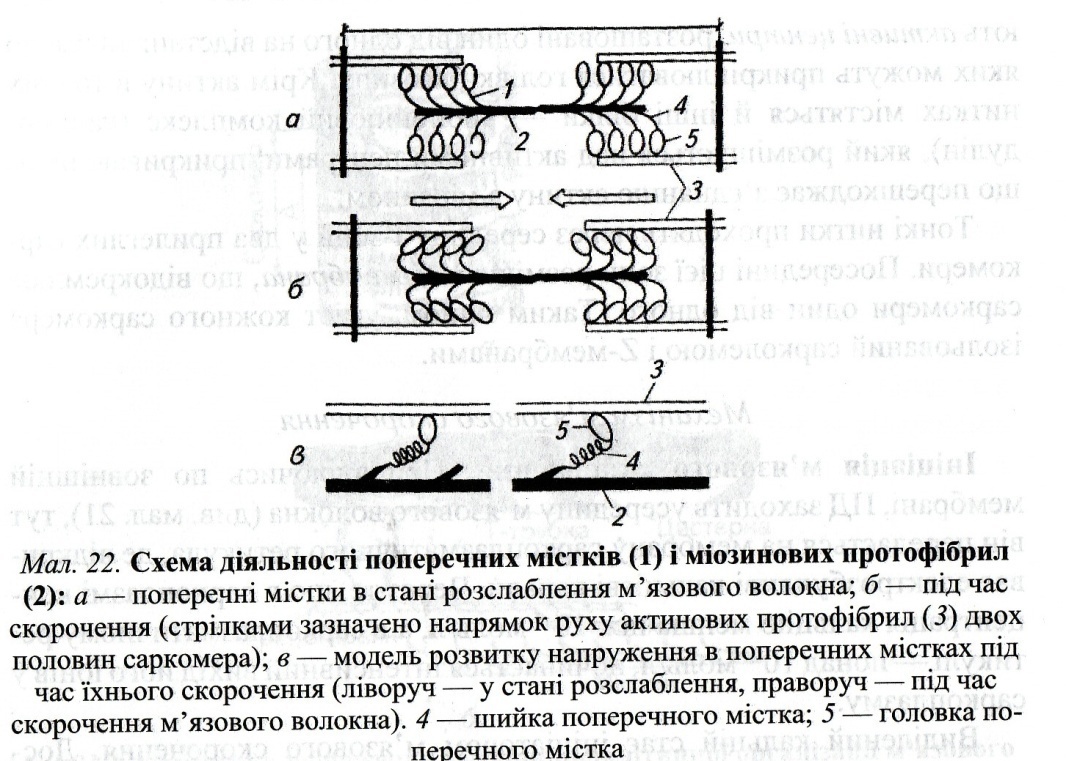
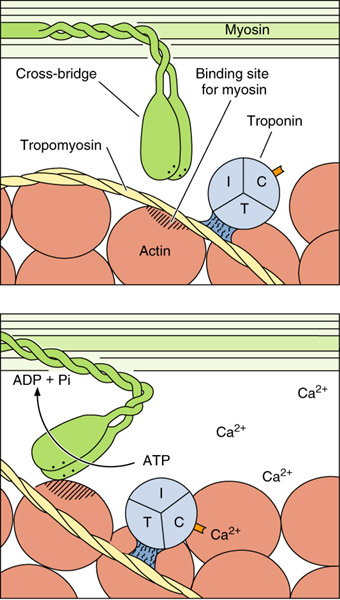
ЗМІСТ ТЕМИ Ініціація м'язового скорочення. Поширюючись по зовнішній мембрані, ПД заходить усередину м'язового волокна, тут він передається на мембрану саркоплазматичного ретикулума, де відкриває електрозбудливі кальцієві канали. Через те, що в саркоплазмі концентрація кальцію менша ніж 10-7моль/л, а в саркоплазматичному ретикулумі - понад 10-4моль/л, починається інтенсивний вихід його іонів у саркоплазму. Виділений кальцій стає ініціатором м'язового скорочення. Достатній для початку м'язового скорочення рівень кальцію досягається через 12-15 мс після приходу нервового імпульсу. Це прихований, латентний, час м'язового скорочення. У зв'язку з тим, що швидкість поширення ПД сарколемою більша від часу, необхідного для виділення Са2+ із саркоплазматичного ретикулума, усі фібрили ділянки м'яза, що іннервуються одним нервом, скорочуються одночасно. В ініціації м'язового скорочення після надходження в саркоплазму Са2+ певну роль відіграє кальмодулін. Приєднуючи Са2+, кальмодулін сприяє активації АТФази й використанню енергії АТФ для зв'язку активного центру актинової нитки з голівкою міозину, а також укороченню м'яза (мал. 22). При з'єднанні кальмодуліну (тропонін С) з кальцієм активний центр актину вивільняється, унаслідок чого до нього приєднується голівка міозину. Ці процеси відбуваються в тому разі, якщо концентрація вільного кальцію в саркоплазмі зростає в 100 і більше разів: з 10-7 до 10-5 моль/л.
" Шарнірний механізм ". Унаслідок поєднання зазначених процесів відбувається: а) підтягування ниток міозину до актинових; б) заряджання міозину енергією, що застосовується для виконання повороту міозинової голівки. Після цього утворювані фосфор й аденозиндифосфорна кислота (АДФ) відходять, а на їхнє місце приєднується нова молекула АТФ, що приводить до розриву зв'язку міозину з активним центром актину. При скороченні м'яза: а) актинові й міозинові нитки практично не коротшають; б) взаємодія актину з міозином призводить до взаємного входження ниток у проміжки між ними; в) дві прилеглі Z-мембрани зближаються одна з одною, і при максимально сильному скороченні відстань між ними може зменшитися майже вдвічі; г) при зменшенні довжини м'яза саркомер розширюється, позаяк укладена всередині саркомера саркоплазма не стискається; ґ) подібні процеси одночасно перебігають у всіх саркомерах м'язового волокна, тому обидва кінці м'яза підтягуються до центру. Нині ще остаточно невідомий механізм, який забезпечує входження актоміозинових ниток одна в одну. Загальноприйнята гіпотеза "шарнірного механізму". Після з'єднання голівки міозину з активним центром актину відбувається її поворот на 45°. Унаслідок розриву містка шийка голівки міозину випрямлюється, набуваючи вихідне положення. За такі рухи ця система й отримала назву шарнірного механізму. Під час повороту міозин просувається актином на один "крок", або "гребок", що дорівнює 20 нм. Надходження нової порції Са2+ приводить до повторення "кроку", але тепер уже іншої голівки, що опинилася навпроти нового активного центру актину, оскільки вони розташовані на відстані близько 40 нм один від одного. У зв'язку з тим що міозинові нитки мають біполярну організацію голівок, то паралельні "гребки" їх забезпечують ковзання актинових ниток вздовж саркомера (від Z-мембрани до його середини).
Розслаблення м'яза. Зазначені процеси ("кроки") повторюватимуться доти, доки саркоплазма міститиме вільний Са2+ (у концентрації понад 10-5 моль/л) й АТФ. Якщо немає нової хвилі деполяризації, кальцій швидко повертається назад у цистерни саркоплазматичного ретикулума. Він відкачується із саркоплазми проти градієнта концентрації за допомогою Са2+-насоса, розташованого на мембрані саркоплазматичного ретикулума. Робота цього насоса, що потребує великої кількості АТФ (для видалення кожного Са2+ використовується 2 молекули АТФ), активується самим кальцієм, точніше, зростанням його концентрації в саркоплазмі. Наслідок відкачування кальцію із саркоплазми - розрив усіх зв'язків актину й міозину і розслаблення м'яза.
Дата добавления: 2015-12-15 | Просмотры: 627 | Нарушение авторских прав |