|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
ОРІЄНТОВНА КАРТКА ДЛЯ РОБОТИ З ЛІТЕРАТУРОЮ.
МАТЕРІАЛИ ДЛЯ САМОКОНТРОЛЮ Дайте відповіді на запитання: 1. Йони якого хімічного елементу ініціюють м’язове скорочення? 2. Назвіть етапи «шарнірного механізму» входження актоміозинових ниток одна в одну. 3. Як відбувається процес розслаблення м’яза? Тема: «Будова та функції нервово-м'язового синапсу»
АКТУАЛЬНІСТЬ ТЕМИ Скороченню скелетних м'язів обов'язково має передувати збудження (потенціал дії), що надходить до них із ЦНС нервовими волокнами. Але мембрана нервового волокна прямого контакту з м'язовою мембраною не має, і передача збудливого імпульсу відбувається за допомогою синапсу.
ФОРМУВАННЯ НАВЧАЛЬНИХ ЦІЛЕЙ ЗНАТИ будову синапсу, механізм синаптичної передачі збудження, причини виникнення синаптичної затримки проведення збудження. ВМІТИ пояснювати шляхи відновлення медіатора у синаптичній бляшці.
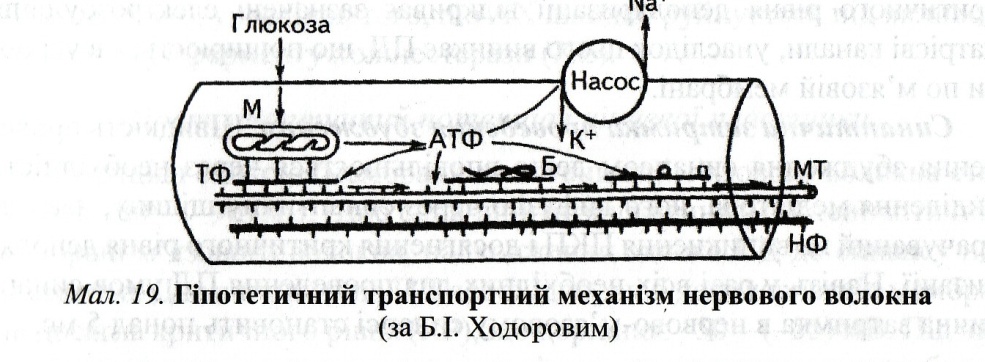
ЗМІСТ ТЕМИ Нервово-м'язовий синапс має таку будову. Нервове волокно закінчується пресинаптичним розширенням. Усередині нього міститься велика кількість (до 300 тис.) синаптичних пухирців (Ø ≈ 50 нм), що містять певну кількість хімічної сполуки - ацетилхоліну (АХ). Це хімічний передавач збудження, що має назву "медіатор". Кожний пухирець містить "квант" медіатора (≈104 молекул АХ). Він передає збудження через синаптичну щілину (завширшки 20-30 нм), що відокремлює дві мембрани: пресинаптичну мембрану нервового закінчення, звернену всередину синапсу, і постсинаптичну - частину мембрани м'язового волокна, що ще називають кінцевою пластинкою. Синаптична щілина забезпечує передачу нервового імпульсу лише в одному напрямку. Синаптична щілина заповнена синаптичною рідиною, що за своїм складом нагадує плазму крові. Пре- і постсинаптична мембрани з'єднуються тонкими ретикулярними волокнами. Крім того, у синаптичній бляшці міститься велика кількість мітохондрій, що свідчить про метаболічну активність цього відділу нервового волокна. Механізм синаптичної передачі збудження. Потенціал дії (ПД), що надійшов до синапсу, просуваючись пресинаптичною мембраною, відкриває електрозбуджувальні кальцієві канали. Са2+ надходить усередину синаптичної бляшки по градієнту концентрації. Збільшення вмісту вільного кальцію (приблизно в 100 разів) забезпечує просування кількох пухирців до пресинаптичної мембрани. Потім мембрана пухирців зливається з мембраною нервового закінчення, і медіатор унаслідок секреції викидається всинаптичну щілину. Медіатор дифундує по синаптичній рідині, і більша частина молекул його досягає постсинаптичної мембрани, де містяться рецептори - холінорецептори (ХР). Зазвичай для передачі одного ПД вивільняється до мільйона молекул АХ (200-300 пухирців). Результат взаємодії АХ із ХР - відкриття хемозбуджувальних іонних каналів. Крізь них можуть проходити лише позитивні іони (стінка каналу електронегативна) натрію і кальцію. Але в нормі, як правило, превалює потік іонів натрію. Вони по концентраційному градієнту із синаптичної щілини надходять усередину м'язового волокна й деполяризують постсинаптичну мембрану. Зазвичай у відкритому стані канал перебуває дуже короткий час (близько 1 мс), унаслідок того, що АХ швидко руйнується під впливом наявного тут ферменту холінестерази (ХЕ). Постсинаптичний потенціал кінцевої пластинки. Потенціал спокою (мембранний потенціал) м'язового волокна становить -80…-90 мВ. Щоб спричинити збудження в постсинаптичній мембрані м'язового волокна, надходження одного ПД до синапсу недостатньо. Для виникнення збудження необхідно, щоб деполяризація постсинаптичної мембрани досягла критичного рівня, що дорівнює -50…-55 мВ. Під час надходження поодинокого кванта медіатора постсинаптична мембрана деполяризується лише на 0,1-0,15 мВ. Різновид такої деполяризації мембрани називають потенціалом кінцевої пластинки (ПКП). ПКП поширюється на прилеглі ділянки мембрани з декрементом, тобто з поступовим зменшенням амплітуди. Унаслідок цього на відносно велику відстань він проводитися не може. Тому ПКП належить до так званих місцевих потенціалів. Важлива його властивість - здатність до сумації. Величина ПКП цілком залежить від кількості медіатора, що виділився. Але якщо під впливом поодинокого ПД виділяється приблизно однакова кількість медіатора, то надходження нервових імпульсів з невеликими проміжками (ПД по нерву можуть передаватися з максимальною частотою до 1 тис. за 1 с) приводить до "викиду" подальших порцій медіатора. Унаслідок того, що рецептори від попереднього АХ вивільняються дуже швидко (через 1-1,5 мс), нове виділення медіатора спричинює повторне відкриття іонних каналів. Новий ПКП, що виникає, нашаровується на ще не зниклу попередню деполяризацію і, підсумовуючись, збільшує амплітуду попереднього ПКП. Виникнення потенціалу дії на мембрані м'язового волокна. Підсумований ПКП при досягненні критичного рівня деполяризації відкриває зазначені електрозбудливі натрієві канали, унаслідок цього виникає ПД, що поширюється в усі боки по м'язовій мембрані. Синаптична затримка проведення збудження. Швидкість проведення збудження синапсом дещо вповільнюється через необхідність виділення медіатора, його дифузію через синаптичну щілину, час, затрачуваний на виникнення ПКП і досягнення критичного рівня деполяризації. Навіть у разі всіх необхідних для проведення ПД умов синаптична затримка в нервово-м'язовому синапсі становить понад 5 мс. Відновлення медіатора в синаптичній бляшці. У зв'язку з тим, що для формування одного ПД на мембрані м'язового волокна потрібен викид приблизно 300 пухирців з АХ, а їх усього близько 300 тис., то через такий синапс могло б пройти лише до 1000 імпульсів. Як правило, скелетні м'язи людини виконують роботу в такому режимі, що цієї кількості ПД явно недостатньо, тому в нервовому волокні відбувається постійне поповнення медіатора. Існує кілька механізмів відновлення пухирців з медіатором. Незабаром після взаємодії АХ з ХР під впливом убудованого в мембрану ферменту ХЕ медіатор руйнується на холін й оцтову кислоту. Більша частина продуктів гідролізу АХ повертається в синаптичну бляшку, де бере участь у ресинтезі нових молекул медіатора, що надходить у знов сформовані пухирці. Ще один метод відновлення витраченого медіатора - активні процеси місцевого синтезу АХ з інших сировинних джерел за допомогою відповідних ферментів, наявних у пресинаптичному закінченні. Транспортна функція нервового волокна. Однак зазначених вище механізмів відновлення медіатора для забезпечення тривалої й інтенсивної м'язової роботи може бути недостатньо. Тому є третій шлях: "підвезення" медіатора від тіла нейрона - аксонний транспорт. У тілі нейрона постійно відбуваються активні процеси синтезу низки сполук, необхідних не лише власне нейрону, а і його відросткам і синапсам. Тут синтезуються різні структурні білки, синаптичні медіатори й низькомолекулярні фактори. Усі вони спускаються до нервового закінчення відростками. Транспортуються й суцільні клітинні органели, зокрема мітохондрії. Аксонний транспорт - активний процес, що використовує енергію АТФ і спеціальні транспортери (мал. 19). Швидкість транспортування речовин по аксону різна. Швидкий транспорт (15-20 мм/год) майже не залежить від розміру сполуки, що транспортується, і виду нейрона. Повільніше транспортуються на деяких ділянках нервового волокна субклітинні структури, наприклад мітохондрії - за добу через поперечний переріз середнього діаметра проходить близько 1000 мітохондрій. Важливою структурною одиницею аксона вважають мікротрубочки. Стінка їх складається з білка тубуліну, а серед нейрофібрил є скоротливий білок актин. Вважають, що актин ковзає уздовж мікротрубочок і "на собі" транспортує субстрат. Цей процес здійснюється із застосуванням енергії АТФ.
Швидкий аксонний транспорт може брати участь і в патологічних процесах. Деякі нейротропні віруси (наприклад, віруси герпесу або поліомієліту) проникають в аксон на периферії і рухаються за допомогою ретроградного транспорту до тіла нейрона, де розмножуються і спричиняють токсичну дію. Токсин правця – білок, який продукується бактеріями, що потрапляють в організм при ушкодженнях шкіри, захоплюється нервовими закінченнями і транспортується до тіла нейрона, блокуючи його, що приводить до розвитку тривалих тонічних судом скелетної мускулатури і різкого підвищення рефлекторної збудливості нейронів, а також до загострення слухової і зорової чутливості. Стомлення нервово-м'язового синапса. Працездатність, тобто здатність проводити збудження, у синапса значно нижча, ніж у нервового волокна. Це може бути наслідком двох явищ. Депресія передачі збудження в синапсі може спричинюватися виснаженням значної частини медіатора або ослабленням його відновлення при занадто високій частоті ПД, що надходять нервовим волокном. Крім того, при інтенсивній м'язовій діяльності недоокиснені продукти (активно утворюються під час гліколізу) знижують чутливість постсинаптичної мембрани до медіатора АХ. Це приводить до зниження амплітуди кожного ПКП і при надмірному зниженні виникнення ПД взагалі стає неможливим.
Дата добавления: 2015-12-15 | Просмотры: 708 | Нарушение авторских прав |