|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Процес сечоутворення. Утворення кінцевої сечі є результатом трьох послідовних процесів (рисУтворення кінцевої сечі є результатом трьох послідовних процесів (рис. 87). Фільтрація Реабсорбція Секреція Фільтрація Фільтрація і реабсорбція і секреція
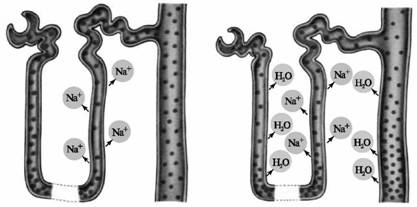
Інулін Глюкоза Іони К+ Сечовина, Діодраст (в нормі) глюкоза (при гіперглікемії) Рис. 87. Процеси, що відбуваються в канальцях при проходженні по них різних компонентів сечі I. У клубочках нефрона відбувається початковий етап сечоутворен-ня—клубочкова фільтрація, ультрафільтрація безбілкової рідини з плазми крові в капсулу ниркового клубочка, у результаті чого утворюється первинна сеча. II. Канальцева реабсорбція — процес зворотного всмоктування речовин, що профільтрувалися, і води з просвіту канальців у кров капілярів нирок. III. Секреція. Клітини деяких відділів канальця переносять з позаклітинної рідини в просвіт нефрона (секретирують) низку органічних і неорганічних речовин або виділяють у просвіт канальця і капіляри речовини, синтезовані в клітині канальця. 15.4.1. Клубочкова фільтрація Ультрафільтрація води і низькомолекулярних компонентів із плазми крові відбувається через клубочковий фільтр. Цей фільтраційний бар'єр майже непроникний для високомолекулярних речовин. Процес ультрафільтрації обумовлений різницею між гідростатичним тиском крові, гідростатичним тиском у капсулі клубочка й онкотичним тиском плазми крові. Фільтруюча мембрана (фільтраційний бар'єр), через яку проходить рідина з просвіту капіляра в порожнину капсули клубочка, складається з трьох шарів: ендотеліальних клітин капілярів, базальної мембрани й епітеліальних клітин вісцерального (внутрішнього) листка капсули — подоцитів. Клітини ендотелія стоншені, товщина цитоплазми бічних частин клітини менша 50 нм. У мембрані є круглі або овальні отвори (пори) розміром 50—100 нм, що займають до 30 \% поверхні клітини. При нормальному кровотоці найбільш великі білкові молекули утворюють шар на поверхні пор і ускладнюють рух через них формених елементів крові і білків. Інші компоненти плазми крові і вода можуть вільно досягати базальної мембрани. Базальна мембрана є однією з найважливіших складових частин фільтруючої мембрани клубочка. У людини товщина базальної мембрани 250—400 нм. Ця мембрана складається з трьох шарів: центрального і двох периферичних. Пори в базальній мембрані перешкоджають проходженню молекул діаметром більше 6 нм. Важливу роль у визначенні розміру фільтрованих речовин відіграють щілинні мембрани між «ніжками» подоцитів. Ці епітеліальні клітини звернені в просвіт капсули ниркового клубочка і мають відростки— «ніжки», якими прикріплюються до базальної мембрани. Базальна мембрана і щілинні мембрани між цими «ніжками» обмежують фільтрацію речовин, діаметр молекул яких більше 6,4 нм. Тому в просвіт нефрона вільно проникає інулін (радіус молекули 1,48 нм, молекулярна маса близько 5200), фільтрується лише 22 \% яєчного альбуміну (радіус молекули 2,85 нм, молекулярна маса 43 500), 3 \% гемоглобіну (радіус молекули 3,25 нм, молекулярна маса 68 000) і менше 1 \% сироваткового альбуміну (радіус молекули 3,55 нм, молекулярна маса 69 000). Проходженню білків через клубочковий фільтр перешкоджають негативно заряджені молекули, що входять до складу базальної мембрани. Обмеження для фільтрації білків, що мають негативний заряд, обумовлено розмірами пор клубочкового фільтра і їх електронеГативністю. Таким чином, склад клубочкового фільтрату залежить від властивостей епітеліального бар'єра і базальної мембрани. Рівень клубочкової фільтрації залежить від різниці між гідростатичним тиском крові (близько 44—47 мм рт. ст. у капілярах клубочка), онкотичним тиском білків плазми крові (близько 25 мм рт. ст.) і гідростатичним тиском у капсулі клубочка (близько 10 мм рт. ст.). Ефективний фільтраційний тиск, що визначає швидкість клубочкової фільтрації, складає 10—15 мм рт. ст. Фільтрація відбувається лише в тому випадку, якщо тиск крові в капілярах клубочків перевищує суму онкотичного тиску білків у плазмі і тиску рідини в капсулі клубочка. Ультрафільтрат у порожнині клубочка практично не містить білків, але подібний плазмі за загальною концентрацією осмотично активних речовин: глюкози, сечовини, сечової кислоти, креатиніну й інших, отже, для розрахунку кількості фільтрованих речовин у клубочках необхідно враховувати, яка їхня частина може проходити з плазми в просвіт нефрона через гломерулярний фільтр. Як будь-який процес пасивного переходу речовин, фільтрація залежить від декількох факторів. Фшьтраіія=рільтр^ а де Рфщщі — фільтраційний тиск; £ — площа фільтрації (в обох нирках близько 1,5—2,0 м2, тобто дорівнює площі поверхні тіла); а—товщина фільтраційної мембрани. Фільтраційний тиск (Рф^ьтр) — це результат дії тиску крові в судинному клубочку, що дарівнює близько 50 мм рт. ст. (Ргідр), онкотичного тиску, створюваного білками плазми крові, що заважає виходу рідини із судини в порожнину капсули, становить близько 30 мм рт. ст. (Ронкотич), тиску фільтрату в капсулі клубочка близько 10 мм рт. ст. (Р ф.). - 1 г * •> г гідр. фільтр7 Рф. = Р — Р — Р. ф. = (50 — 30 — 10) = 10 мм рт. ст. фільтр гідр онкотич гідр. фільтр 4 / г Вимірювання швидкості клубочкової фільтрації Величина зміни швидкості фільтрації оцінюється по тому, як йде з плазми речови-на-маркер. Як міра цього переходу використовується коефіцієнт очищення — кліренс (від англ. «clear» — чистий, світлий). Вимоги до речовини-маркера, кліренс якого дозволяє оцінити величину фільтрації: — нетоксичність речовини; — хороша його розчинність у воді; — нездатність зв'язуватися з білками плазми або фільтраційної мембрани; — здатність фільтруватися; — нездатність реабсорбуватися і секретируватися. Як такі речовини частіше використовують полімер фруктози — інулін. Отже, як показник фільтрації використовують кліренс, тобто оцінюють, скільки плазми очистилося від інуліну в одиницю часу, тобто профільтрувалося. Розглянемо принцип очищення на прикладі виміру об'єму клубочкової фільтрації за допомогою інуліну. Визначення проводять на тлі постійної концентрації інуліну в крові. Для цього або постійно вводять цей полімер під час дослідження крові, або вводять його в надлишку. Кількість інуліну, що профільтрувався в клубочках, (In) фільтрату добутку об'єму фільтрату (CIn) на концентрацію в ньому інуліну (вона дорівнює його концентрації в плазмі крові, РІп). Кількість інуліну, що виділилася за той же час із сечею дорівнює добутку об'єму екскретированної сечі (V) на концентрацію в ній інуліну (Ub). Тому що інулін не реабсорбується і не секретирується, то кількість інуліну, що профільтрувався, (Сы- РІп) дорівнює кількості, що виділилась (V-UIn), звідки: СІп= UIn'V/ PIn. Ця формула є основою для розрахунку швидкості клубочкової фільтрації. Швидкість фільтрації рідини обчислюють у мл/хв; для зіставлення величини клубочкової фільтрації в людей різних маси тіла і росту її відносять до стандартної поверхні тіла людини (1,73 м2). У нормі в чоловіків в обох нирках швидкість клубочкової фільтрації на 1,73 м2 складає близько 125 мл/хв, у жінок — приблизно 110 мл/хв. У здорової людини вода попадає в просвіт нефрона в результаті фільтрації в клубочках, реабсорбується в канальцях, і внаслідок цього концентрація інуліну зростає. Концентраційний показник інуліну UIn/PIn вказує, у скільки разів зменшується об'єму фільтрату при його проходженні по канальцях. Ця величина має важливе значення для судження про особливості обробки будь-якої речовини в канальцях, для відповіді на питання про те, чи піддається речовина реабсорбції чи секретирується клітинами ка-нальців. Якщо концентраційний показник даної речовини Х U /Рх менший, ніж одночасно обмірювана величина UIn /PIn, то це вказує на реабсорбцію речовини Хв канальцях. Якщо U /Рх більше, ніж UIn/PIn, то це вказує на його секрецію. Відношення концентраційних показників речовини Хта інуліну U /Рх: UIn /PIn називається екскреторною фракцією (EF). Фактори, що впливають на величину фільтрації. Тому що Рфільтр залежить від Ргідр, Ронк, Ргідр фільтр, то будь-яка їхня зміна позначається на фільтрації і, у підсумку, на величині діурезу. Розглянемо випадок припинення діурезу. Це може мати місце при: зниженні Ргідр при великій крововтраті, різкому зниженні загального тиску крові (колапс), звуженні судин у судинному клубочку через біль; ріст Ронк при підвищенні вмісту білків у плазмі, зневоднюванні організму при спразі, сильному потінні, блювоті, проносі; ріст Ргідр фільтр (неможливість відтоку з нирки при ниркових каменях, звуженнях сечовивідних шляхів при запаленнях, при пухлинах, що здавлює сечоводи, при їхньому перегині при зміщенні нирки). 15.4.2. Канальцева реабсорбція Більша частина первинної сечі повертається в кров за рахунок механізму реабсорбції (зворотного всмоктування). І лише нереабсорбо-вана її частина виділяється як кінцева сеча. Так, за 1 хв фільтрується 100—140 мл плазми (за добу до 180 л). Виділяється у виді сечі 1—2 мл/хв (за добу1—1,5 л). Різниця (180 л—1,5 л) і складає величину реабсорбції за добу. У нормі реабсорбується 94—99 \% первинної сечі. Роль клітин у різних сеґментах нефрона в реабсорбції неоднакова. З'ясовано особливості реабсорбції різних речовин у різних частинах ниркових канальців. У проксимальному сеґменті нефрона практично цілком реабсорбуються амінокислоти, глюкоза, вітаміни, білки, мікроелементи, значна кількість іонів Na+, Cl—, HCO3—. У результаті активного поглинання Na+ і супровідних його аніонів осмотичний тиск фільтрату знижується й у капіляри шляхом осмосу переходить еквівалентна кількість води. У підсумку, у канальці утворюється фільтрат, ізотонічний крові капілярів. Цей фільтрат надходить у петлю Генле. Тут відбувається подальша реабсорбція і концентрування сечі за рахунок поворотно-протиточної системи — фізіологічного механізму, основаного на різній проникності стінок низхідної і висхідної ділянок петлі для води і Na+ (рис. 88). Концентрування сечі відбувається в такий спосіб. У висхідній частині петлі нефрона реабсорбуються Na, К, Са, Mg, C1, сечовина. Переходячи в міжклітинну рідину, вони підвищують її осмотичний тиск, тому що Н2О в цьому відділі не реабсор-бується. Низхідна частина петлі Генле пропускає воду, що рухається в міжклітинному просторі за законами осмосу, тому що осмотичний тиск там вище за рахунок виходу іонів Na+, С1—, К+ з висхідної частини. Вихід Н2О із низхідної частини петлі приводить до того, що сеча стає більш концентрованою стосовно плазми крові. Це сприяє реабсорбції Na+ у висхідній частині петлі, що, у свою чергу, викликає вихід H2O унизхідній частині. Ці два процеси сполучені, у підсумку сеча втрачає в петлі Генле велику кількість Н2О і Na+, тобто стає більш концентрованою. Таким чином, роль петлі Генле як протиточного концентруючого механізму визначають такі фактори: 1. Близьке сусідство низхідного і висхідного коліна. 2. Проникність низхідного коліна для Н2О. 3. Непроникність низхідного коліна для розчинених речовин. 4. Проникність висхідного сеґмента для №+, К+, Са2+, М^2+, С1-. 5. Наявність механізмів активного транспорту у висхідному коліні.
Реабсорбція № Реабсорбція Н2О слідом за № Рис. 88. Діяльність протиточного механізму в нирках. Послідовні етапи виходу №+ і води із канальнії! У дистальній частині канальця, що підходить назад до мальпігієво-го тільця, відбувається подальша реабсорбція №+, К+, Са2+, Mg2+, Н2О, що залежить від концентрації цих речовин у крові — факультативна реабсорбція. Якщо їх багато, то вони не реабсорбуються, якщо мало, то вони повертаються в кров. Дистальний відділ реґулює і підтримує сталість концентрації іонів №+ і К+ в організмі. Проникність стінок дистальної частини канальця для Н2О регулюється антидуретичним гормоном гіпофіза (секреція якого залежить від осмотичного тиску крові). При підвищенні осмотичного тиску (тобто при зменшенні кількості Н2О) секреція вазопресину збільшується, збільшується проникність стінок канальця для Н2О, і вона затримується в організмі. У такий же спосіб регулюється реабсорбція води в збірній трубочці, що також бере участь в утворенні гіпертонічної чи гіпотонічної сечі, у залежності від потреби організму у воді. Реабсорбція натрію і хлору являє собою найбільш значний за об'ємом й енергетичними витратами процес. У проксимальному ка нальці в результаті реабсорбції більшості речовин і води, що профільтрувалися, об'єм первинної сечі зменшується, і в початковий відділ петлі нефрона надходить близько 1/3 профільтрованої в клубочках рідини. З усієї кількості натрію, що надійшов у нефрон при фільтрації, у петлі нефрона всмоктується до 25 \%, у дистальному звивистому канальці — близько 9 \%, і менше 1 \% реабсорбується в збірних трубочках або екс-кретирується із сечею. Реабсорбція в дистальному сеґменті характеризується тим, що клітини переносять меншу, ніж у проксимальному канальці, кількість іонів, але проти більшого градієнта концентрації. Цей сеґмент нефрона і збірні трубки відіграють найважливішу роль у регуляції об'єму виділюваної сечі і концентрації в ній осмотично активних речовин. У кінцевій сечі концентрація натрію може знижуватися до 1 ммоль/л у порівнянні з 140 ммоль/л у плазмі крові. Удистальному канальці калій не тільки реабсорбується, але і секретирується при його надлишку в організмі. У проксимальному ві дділі нефрона реабсорбція натрію, калію, хлору й інших речовин відбувається через високопроникну для води мембрану стінки канальця. Навпаки, у висхідному відділі петлі нефрона, дистальних звивистих канальцях і збірних трубках реабсорбція іонів і води відбувається через малопроникнудля води стінку канальця. Проникність мембрани для води в окремих ділянках нефрона і збірних трубках може регулюватися, а величина проникності змінюється в залежності від функціонального стану організму. Під впливом імпульсів, що надходять по еферентних нервах, і при дії біологічно активних речовин реабсорбція натрію і хлору регулюється в проксимальному відділі нефрона. У випадку збільшення об'єму крові і позаклітинної рідини, коли зменшення реабсорбції в проксимальному канальці сприяє посиленню екскреції іонів і води і тим самим — відновленню водно-сольової рівноваги, у проксимальному канальці завжди зберігається ізоосмія. Стінка канальця проникна для води, і об'єм реабсорбованої води визначається кількістю реабсорбованих осмотично активних речовин, за якими вода рухається по осмотичному градієнті. У кінцевих частинах дистального сегмента нефрона і збірних трубках проникність стінки канальця для води регулюється вазопресином. 15.4.2.1. Механізми канальцевої реабсорбції' Зворотне всмоктування різних речовин у канальцях забезпечується активним і пасивним транспортом. Якщо речовина реабсорбується проти електрохімічного і концентраційного градієнтів, процес нази вається активним транспортом. Розрізняють два види активного транспорту — первинно-активний і вторинно-активний. Первинно-активним транспорт називається в тому випадку, коли відбувається перенесення речовини проти електрохімічного градієнта за рахунок енергії клітинного метаболізму. Прикладом служить транспорт іонів №+, що відбувається при участі ферменту №+, К+-АТФази, що використовує енергію АТФ. Вторинно-активним називається перенесення речовини проти концентраційного градієнта, але без витрати енергії клітини безпосередньо на цей процес: так реабсорбуються глюкоза, амінокислоти. З просвіту канальця ці органічні речовини надходять у клітини проксимального канальця за допомогою спеціального переносника, що обов'язково повинний приєднати іон №+. Цей комплекс (переносник + + органічна речовина+№+) сприяє переміщенню речовини через мембрану і надходженню його усередину клітини. Рушійної силою перенесення цих речовин через плазматичну мембрану служить менша в порівнянні з просвітом канальця концентрація натрію в цитоплазмі клітини. Ґрадієнт концентрації натрію обумовлений безперестанним активним виведенням натрію з клітини в позаклітинну рідину за допомогою №+, К+-АТФази, локалізованої в мембрані клітини. Мембрана клітин канальців, повернена в просвіт канальця, називається апікальною, а спрямована у бік міжклітинної речовини — базальною. Реабсорбція води, хлору і деяких інших іонів, сечовини здійснюється за допомогою пасивного транспорту — по електрохімічному, концентраційному чи осмотичному градієнту. Прикладом пасивного транспорту є реабсорбція в дистальному звивистому канальці хлору по електрохімічному градієнту, створеному активним транспортом натрію. По осмотичному градієнту транспортується вода, причому швидкість її всмоктування залежить від осмотичної проникності стінки канальця і різниці концентрації осмотично активних речовин по обидва боки його стінки. У вмісті проксимального канальця внаслідок всмоктування води і розчинених у ній речовин росте концентрація сечовини, невелика кількість якої по концентраційному градієнту реабсорбується в кров. Клітинний механізм реабсорбції іонів розглянемо на прикладі №+. У проксимальному канальці нефрона всмоктування №+ у кров відбувається в результаті ряду процесів, один із яких — активний транспорт №+ із просвіту канальця, інший — пасивна реабсорбція №+ слідом за іонами гідрокарбонату та СІ-, що транспортуються активно в кров. При введенні одного мікроелектрода в просвіт канальців, а другого в нав-колоканальцеву рідину було виявлено, що різниця потенціалів між зовнішньою і внутрішньою поверхнею стінки проксимального канальця може досягати — 60 мВ. Просвіт обох канальців електронегативний, а в крові (отже, і в позаклітинній рідині) концентрація №+ вища, ніж у рідині, що знаходиться в просвіті канальців. Тому реабсорбція здійснюється активно проти градієнта електрохімічного потенціалу. При цьому з просвіту канальця входить у клітину по натрієвому каналу або за участю переносника. Внутрішня частина клітини заряджена негативно, і позитивно заряджений №+ надходить у клітину по градієнту потенціалу, рухається у бік базальної плазматичної мембрани, через яку натрієвим насосом викидається в міжклітинну рідину; градієнт потенціалу на цій мембрані досягає 70—90 мВ. Фільтрована глюкоза практично цілком реабсорбується клітинами проксимального канальця, і в нормі за добу із сечею виділяється незначна її кількість (не більш 130 мг). Процес зворотного всмоктування глюкози здійснюється проти високого концентраційного градієнта і є вторинно-активним. В апікальній мембрані клітини глюкоза з'єднується з переносником, що повинний приєднати також №+, після чого комплекс транспортується через мембрану, тобто в цитоплазму надходять глюкоза і №+. Апікальна мембрана відрізняється високою селективністю й однобічною проникністю і не пропускає ні глюкозу, ні №+ назад із клітини в просвіт канальця. Перенесення глюкози з клітини в кров через базальну плазматичну мембрану носить характер полегшеної дифузії, а №+, як уже зазначалося вище, видаляється натрієвим насосом, що знаходиться в цій мембрані. Амінокислоти майже цілком реабсорбуються клітинами проксимального канальця. Існує не менше чотирьох систем транспорту амінокислот із просвіту канальця в кров. Кожна з цих систем забезпечує всмоктування низки амінокислот однієї групи. Так, система реабсорбції двоосновних амінокислот бере участь у всмоктуванні лізину, аргініну, орнітину і, можливо, цистину. При введенні в кров надлишку однієї з цих амінокислот починається посилена екскреція ниркою амінокислот лише даної групи. Виділення із сечею слабких кислот і основ залежить від їхньої клу-бочкової фільтрації, процесу реабсорбції чи секреції. Процес виведення цих речовин багато в чому визначається «неіонною дифузією», вплив якої особливо виявляється в дистальних канальцях і збірних трубках. Невелика кількість білка, що профільтрувався в клубочках, реабсорбується клітинами проксимальних канальців. Виділення білків із сечею в нормі складає не більше 20—75 мг на добу, а при захворюваннях нирок воно може зростати до 50 м на добу. Збільшення виділення білків із сечею (протеінурія) може бути обумовлена порушенням їхньої реабсорбції або збільшенням фільтрації. На відміну від реабсорбції електролітів, глюкози й амінокислот, які, проникнувши через апікальну мембрану, у незміненому виді досягають базальної плазматичної мембрани і транспортуються в кров, реабсорбція білка забезпечується принципово іншим механізмом: білок попадає в клітину за допомогою піноцитозу. Молекули білка, що профільтрувався, адсорбуються на поверхні апікальної мембрани клітини, при цьому мембрана бере участь в утворенні піноцитозного пухирця. Цей пухирець рухається у бік базальної частини клітини. В навколоядерній області, де локалізований пластинчастий комплекс (апарат Гольджі), пухирці зливаються з лізосомами, що мають високу ферментативну активність. У лізосомах захоплені білки розщеплюються й амінокислоти, що утворилися, відходять в кров через базальну плазматичну мембрану. Варто підкреслити, що не усі білки піддаються гідролізу в процесі транспорту і частина їх переноситься в кров у незміненому вигляді. Сечовина. У людини і більшості хребетних азот, що утворюється при розпаді білків, екскретирується в основному як сечовина. Ця дрібна електронейтральна молекула вільно фільтрується, але одночасно так легко дифундує, що в проксимальному канальці близько 1/3 сечовини повертається в кров. У дистальній частині нефрона й у кінцевому відділі збірної трубочки дифузійна проникність для сечовини низька, але тут вона може реабсорбуватися по механізму проходження за розчинником. Тому, якщо в дистальних відділах всмоктується велика кількість води й утворюється сеча із високою осмотичною концентрацією, разом з водою повертається в кров майже третина відфільтрованої сечовини. У випадку водяного діурезу всмоктування води в дистальному канальці припиняється, і сечовини відповідно виводиться більше. Таким чином, її екскреція залежить від діурезу. Через фільтр вона проходить з такою ж швидкістю, як індикаторна речовина інулін. Однак її кліренс завжди менший, принаймні, на 1/3, оскільки саме ця частка реабсорбується в проксимальному канальці незалежно від діурезу. Ще 1/3 може реабсорбуватися в дистальних відділах нефрона. Сечова кислота. У вигляді сечової кислоти в людини виводиться тільки 5 \% азоту. Однак ця сполука має велике клінічне значення, оскільки пов'язана з виникненням подагри й утворенням ниркових каменів — захворюваннями, що набувають в останні десятиліття все більшого поширення. Сечова кислота—кінцевий продукт обміну пуринів. Сечова кислота вільно фільтрується. Як й інші органічні кислоти, вона також дістається з навколоканальцевої крові клітинами проксимальних канальців і секретирується в канальцеву рідину. Проте одночасно вона тут же реабсорбується, тому наприкінці цього відділу її при близно стільки ж, скільки початково відфільтрувалось. У низхідному коліні петлі Генле переважає реабсорбція, і до вершини цієї структури доходить лише близько 10 \% сечової кислоти. Саме ця кількість виділяється із сечею, тому що вона пройде через відділи нефрона, розташовані в нирковому сосочку, важливо з двох причин: її розчинність у воді, по-перше, обмежена, а по-друге, залежить від рН. рКа сечової кислоти дорівнює 5,8, тому при нормальному рН крові (7,4) вона майже цілком дисоційована до урат-аніона. Дисоційована до урату, сечова кислота в 20 разів більш розчинна, ніж у недисоційованій формі. Проте канальцева рідина, проходячи через дистальний відділ нефрона, зменшується в об'ємі і продовжує підкислюватися, тому концентрація сечової кислоти зростає, а її дисоціація пригнічується. У збірній трубочці, де рН дорівнює 5,8, вона на 50 \% не дисоційована; при рН 4,5 ця частка зростає до 95 \%. 15.4.3. Канальцева секреція У виділенні продуктів обміну і чужорідних речовин має значення їх секреція з крові в просвіт канальця проти концентраційного й електрохімічного градієнтів. Цей механізм виділення речовин дозволяє швидко екскретирувати деякі органічні кислоти і основи, а також деякі іони, наприклад К+. Секреція органічних кислот (феноловий червоний, ПАГ, діодраст, пеніцилін) і органічних основ (холін) відбувається в проксимальному сегменті нефрона й обумовлена функціонуванням спеціальних систем транспорту. Калій секретирується в кінцевих частинах дистального сегмента і збірних трубок. Розглянемо механізм процесу секреції органічних кислот на прикладі виділення ниркою ПАГ (парааміногіпурат). При введенні ПАГ у кров людини її виділення із сечею залежить від фільтрації в клубочках і секреції клітинами канальців. Коли секреція ПАГ досягає максимального рівня (ТтРАН), вона стає постійною і не залежить від вмісту ПАГ у плазмі крові. Принцип секреторного процесу при транспорті органічних сполук полягає в тому, що в мембрані клітини проксимального канальця, поверненої до інтер-стиціальної рідини, є переносник А, що має високу спорідненість до ПАГ. У присутності ПАГ утворюється комплекс А-ПАГ, який забезпечує переміщення ПАГ через мембрану, і на її внутрішній поверхні ПАГ вивільняється в цитоплазму. При цьому переносник знову набуває здатності переміщатися до зовнішньої поверхні мембрани і з'єднуватися з новою молекулою ПАГ. Механізм транспорту полягає в тому, що переносник обмінює ПАГ на а-кетоглутарат на базальній плазматичній мембрані клітини проксимального канальця. Переносник забезпечує надходження ПАГ усередину клітини. Рівень секреції залежить від кількості переносників у мембрані. Секреція ПАГ зростає пропорційно збільшенню концентрації ПАГ у крові доти, поки всі молекули переносника не насичуються ПАГ. Максимальна швидкість транспорту ПАГ досягається в той момент, коли кількість ПАГ, доступна для транспорту, дорівнює кількості молекул переносника А, що можуть утворювати комплекс А-ПАГ. Надійшовший у клітину ПАГ рухається по цитоплазмі до апікальної мембрани і за допомогою наявного в ній спеціального механізму виділяється в просвіт канальця. Секреція аміаку. Принцип неіонної дифузії лежить в основі виведення аміаку. Джерелом 1ЧН3 є обмін амінокислот і головним чином глутаміну в клітинах епітелію канальців. Аміак має високу розчинність у жирах і легко проникає через мембрану у фільтрат. І якщо його тут не зв'язати, то він так само легко може повернутися в клітину, а потім і в позаклітинну рідину. Але в сечі протікає реакція зв'язування аміаку з Н+, завдяки чому аміак знаходиться в рівноважному стані з амонієм: ічн3 + н+ <=> ічн4. Іон амонію погано проникає через мембрану і, зв'язуючись з катіонами, виділяється із сечею. Таким чином, кисла сеча, що містить багато Н+, сприяє більшій екскреції аміаку. Активна секреція органічних кислот і основ. У проксимальних ка-нальцях є три типи транспортних систем, які активно (із використанням АТФ) секретирують різні речовини. Одна з них секретирує органічні кислоти (парааміногіпурову, сечову кислоти, пеніцилін і т. д.). Друга—секретирує відносно сильні органічні основи (гуанідин, холін), третя — етилендіамінтетраацетат. Функціонують вони незалежно одна від одних. Фільтрація і секреція водню. Участь нирок у підтримці кислотно-основного стану (КОС) зумовлена очищенням крові від нелетучих кислот і основ, що утворюються в процесі метаболізму або надходять у надзвичайно великій кількості з їжею. У процесі розщеплення білків і нуклеїнових кислот утворюються аніони — фосфати і сульфати. Компенсація зрушення рН при цьому відбувається в два етапи. Спочатку вони нейтралізуються шляхом перетворення аніона крові НСО3-у СО2-, який виділяється через легені. Потім аніони сильних кислот виводяться в ниркових канальцях, а НСО3- реабсорбується, що підтримує буферну ємність крові. Ці процеси тісно сполучені з іншими процесами утворення сечі і характеризуються надзвичайною економічністю. Частина іонів виводиться, інші можуть затримуватися чи обмінюватися на ті, які менш важливі для організму. Так, наприклад, необхідний для забезпечення багатьох життєво важливих процесів натрій може обмінюватися на водень або калій. Іони, що виводяться в сечу, змінюють її рН у досить широких межах (від 4,5 до 8,6). При максимальному закисленні сечі іон Н+ виводиться як у вільному стані (за добу до 50 мкмоль Н+), так і в зв'язаному із буферними сполуками. Іони водню в сечу надходять не тільки з фільтратом, вони можуть секретируватися в дистальних і проксималь них відділах канальців нефрона. У проксимальних відділах секреція Н+ відбувається шляхом обміну на при реабсорбції останнього. Обмінна на №+ секреція відбувається й у дистальних відділах, тобто там, де відбувається його реабсорбція, регульована альдостероном. Дата добавления: 2015-12-16 | Просмотры: 1150 | Нарушение авторских прав |