|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Анатомо-гістологічний методПлоща кори великих півкуль складає 0,25 м2. Кількість нейронів (Н) у корі товщиною від 1,5 до 4,5 мм досягає 1´109. У корі 6 шарів (рис. 4.40). 1. Молекулярний (переважно з дендритів пірамідних Н й волокон неспецифічних ядер таламусу – регуляторів збудливості кори). 2. Зовнішній зернистий шар – із зірчастих клітин, через які циркулює збудження, що важливо для пам’яті. 3. Шар малих пірамід (зовнішній пірамідний) – з клітин, які забезпечують внутрішньокоркові зв’язки. 4. Внутрішній зернистий – із зірчастих клітин, на яких закінчуються волокна специфічних таламо- кортикальних шляхів. 5. Шар великих пірамід, які започатковують своїми аксонами кортико-спинальний (пірамідний) тракт. 6. Поліморфний шар, клітини якого започатковують кортико-таламічні, кортико-каудальні та інші міжцентральні зв’язки. Гістохімічно в корі виявлено ГАМК як медіатор, що викликає гальмування. Таким чином було доведено наявність гальмівних коркових нейронів. Електрофізіологічно блискуче визначено декілька видів гальмівних коркових нейронів (Сєрков, 1986). Є також глутамат, як медіатор мотонейронів, причетних до утворення внутрішньокоркових зв’язків. Багато в корі серотоніну. З урахуванням вираженості шарів кори, кількості та виду в них нейронів (їх щільності) у корі описано цитоархітектонічні поля – 53 за Бродманом (рис. 4.41), 266 за Фохтом та Економо. Дослідженнями, започаткованими Монкаслом та Едельменом, установлено колонкову будову кори (рис. 4.42). Колонка – це структурно-функціональна одиниця кори, яка реагує на подразнення певної модальності, а своїми зв’язками закінчується на одному пулі мотонейронів (нагадуємо, що пул – це сукупність мотонейронів, іннервованих одним чутливим нейроном). Розташування пірамідних клітин у напрямку, перпендикулярному до поверхні кори, відповідає розташуванню більшості вставних нейронів, що й утворює гістологічно відособлені мікроколонки з діаметром 80 мкм (діаметр пірамідної клітини складає 50-100 мкм). Вхідний аферентний імпульс проходить по одній з аферентних гілочок у товщу кори, активуючи
Між колонками є зв’язки за рахунок зворотних кола- тералей. Таким чином створюється модуль – функціональна колонка з діаметром 0,3-1 мм. Їх у корі нараховується біля 3.106. У складі модуля розташовано сотні пірамідних клітин зі схожими функціями. Модулі перекривають один одного, серед них є збуджувальні й гальмівні. Отже, модуль регулює діяльність декількох м’язів, але таких, що діють на певний суглоб. Таким чином, у корі представлено не стільки м’язи, скільки окремі рухи. У складі модуля може створюватись декілька варіантів ансамблів, мінливих у часі. Тому ефект при повторних подразненнях може бути неодна- ковим. Крім того, аферентне волокно, дивергуючи, здатне активу- вати досить широку зону – розміром до 0,1 мм. У зоровій корі це до 5000 нейронів. Отже, один чутливий нейрон (його ре- цептор) проекує сигнал на ціле поле нейронів кори з їх кола- тералями й зв’язками. Це забезпечує проекцію не точка в точку, а на багато нейронів, що необхідно для повного аналізу й можливості передачі в інші “зацікавлені” структури. У цьому полягає екранний принцип функціонування коркових полів.
Рис. 4.40. Нейронна будова кори головного мозку. I – молекулярний шар, II – зовнішній зернистий шар, III – шар малих пірамід (зовнішній пірамідний), IV – внутрішній зернистий, V – шар великих пірамід (внутрішній пірамідний), VI – поліморфний шар.
Рис. 4.41. Цитоархітектонічні поля за Бродманом.
Рис. 4.42. Струк- турний модуль сенсомоторної кори (за О.Д.Ноздрачевим, 1991). III – VI – коркові ша- ри; 1 – інтернейрони; 2 – аферентний вхід на інтернейрон; 3 – аферентний вхід на інтернейрон і пірамідний нейрон; 4 – зворотна раль аксона, яка контактує з гальмівним інтернейроном; 5 – пучок аксонів, який виходить за межі колонки; 6 – зворотна колатераль із полегшуючими впливами у межах модуля; 7 – кортикоспінальні пірамідні клітини; 8 – кортикорубральна пірамідна клітина. Дата добавления: 2015-12-15 | Просмотры: 655 | Нарушение авторских прав |
 зірчасті клітини й опрацьовуючись тут з охопленням відповідної великої пірамідної клітини та декількох ма- лих (III шару). Звідси оброблений сигнал покидає кору через волокна пірамідної клітини й надходить до інших структур мозку.
зірчасті клітини й опрацьовуючись тут з охопленням відповідної великої пірамідної клітини та декількох ма- лих (III шару). Звідси оброблений сигнал покидає кору через волокна пірамідної клітини й надходить до інших структур мозку.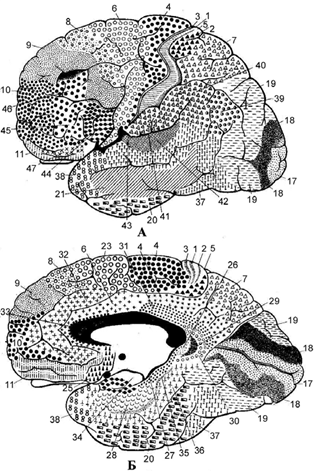
 А – зовнішня поверхня; Б – медіальна поверхня.
А – зовнішня поверхня; Б – медіальна поверхня.