|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Эволюция человекачто механизмы, ответственные за большинство видовых различий между человеком и человекообразными обезьянами, связаны с перицентрическими инверсиями [1929]. Различие в числе хромосом можно объяснить объединением двух различных акроцентрических хромосом, по длине примерно равных D-хромосомам. Такое событие, вероятно, привело к образованию одной длинной субметацентрической хромосомы - 2-й хромосомы человека. Хорошо известно, что объединение разных хромосом имеет место и в современной человеческой популяции: как правило, это происходит при центрическом слиянии, которое сопряжено с потерей коротких плеч хромосомы. Вполне возможно, что именно этот феномен является причиной обнаруженных видовых различий. Детальный анализ результатов дифференциального окрашивания показал, что материал коротких плеч имеется. Хромосома номер 2 человека возникла в результате теломерного слияния [1912]. Подобного рода события приводят к формированию хромосом, имеющих две центромеры (дицентриков), что в свою очередь сопровождается нарушениями митоза, аналогичными тем, которые индуцируются радиацией или являются результатами обменов, следующих за разрывами хромосом (разд. 5.2.1.1). Митоз может идти нормально только в том случае, если функционирует лишь одна из центромер дицентрика. Такой феномен действительно наблюдался в случае хромосомных аберраций, возникающих в настоящее время. Детальный анализ всех визуально обнаружимых перестроек, по которым виды антропоидов отличаются друг от друга и от человека, был проведен Дютрилло (1975) [1912]. Пример. На рис. 7.4 представлены фотографии гомологов 2-й хромосомы человека. Хромосомы Ропgо и Gorilla отличаются инверсией в 2q, a Gorilla и Pan другой инверсией, локализованной в 2р. Теломерное слияние должно было произойти после разделения предков Pan и Homo. На некоторых препаратах 2-й хромосомы человека видна вторичная перетяжка в точке слияния (2qh). Изредка (очень редко) наблюдается эндоредупликация сегментов, соответствующих бывшей 2q-xpoMocoMe, что свидетельствует об известной независимости партнеров, слившихся
друг с другом. На основе этих результатов можно реконструировать ход эволюции 2-й хромосомы. Сравнение кариотипов пяти видов. Межвидовые различия использованы для реконструкции эволюции всех хромосом. Кроме одного теломерного слияния и перицентрических инверсий было выявлено несколько парацентрических инверсий. В табл. 7.3 приведены числа хромосомных перестроек разного типа, по которым отличаются сравниваемые виды. Как и ожидалось, наибольшее сходство обнаруживают два вида шимпанзе. Ближайшим родственником Homo является шимпанзе, а самым далеким - Ропдо. К такому же выводу можно прийти на основании морфологических данных. Противоречия в полученных данных и их возможное объяснение с неортодоксальных позиций. Для каждой отдельной хромосомы можно установить эволюционные взаимосвязи между различными перестройками, имеющимися у разных видов, и выяснить их филогению. Однако, когда эти хромосомные филогении были совмещены друг с другом с целью реконструкции общей фи- 7. Эволюция человека 11
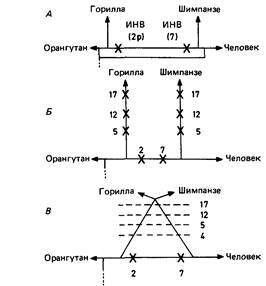
логении, возникло неожиданное затруднение. Оказалось, что Pan troglodytes и Gorilla имеют три общие инверсии (5, 12 и 17), что свидетельствует о наличии у них общего предка, не являвшегося предком Homo; с другой стороны, Homo и Pan имеют две общие инверсии (2, 7), не найденные у Gorilla. Последний результат свидетельствует о существовании общего предка этих двух видов, не являющегося предком Gorilla. Как разрешить эту дилемму? Дютрилло (1975) [1912] предложил три возможных объяснения данного противоречия (рис. 7.5): 1. Общая предковая популяция, возможно, имела хромосомный полиморфизм по этим перестройкам. Однако этот полиморфизм должен был существовать в течение длительного времени, вероятно, на протяжении миллионов лет. Кроме того, мы знаем, что перицентрические инверсии, захватывающие такой большой район хромосомы, как в этом случае, могут приводить к нарушениям мейоза и, следовательно, влиять на репродукцию. Отбор против гетерозигот, который здесь действует, не приводит к стабильному генетическому равновесию (разд. 6.2.1.4). С другой стороны, в Финляндии зафиксирована высокая концентрация перицентрических инверсий, частоты которых почти удовлет-
12 7. Эволюция человека воряют критерию полиморфизма (разд. 2.2.2). 2. Второе объяснение сводится к предположению, что в филогенетических линиях Gorilla и Pan troglodytes произошла независимая фиксация трех идентичных перестроек. Однако это событие маловероятно. Поэтому наиболее правдоподобным Дютрилло считает третье из возможных объяснений. 3. Между популяциями предков Gorilla и Pan, после их отделения от предков человека, время от времени происходила гибридизация. Возможно, что вначале произошло разделение предков Gorilla и общих предков шимпанзе и человека, которое сохранялось, вероятно, благодаря экологической изоляции. Много позже разделились линии шимпанзе и человека; предки Pan вселились в ту же экологическую нишу, в которой уже обитала протогорилла (вероятно, эта ниша приурочена к влажным тропическим лесам). Здесь могла происходить повторная гибридизация, продолжавшаяся до тех пор, пока эти два вида не выработали, наконец, репродуктивные барьеры, чем и завершился процесс видообразования. Другое труднообъяснимое видовое различие-это наличие или отсутствие определенных хромосомных сегментов. Присутствие или отсутствие определенных сегментов. Помимо описанных выше перестроек у некоторых из этих приматов регулярно выявляются определенные хромосомные сегменты, отсутствующие у других видов: а) терминальные Q-сегменты: после окрашивания акрихин-ипритом на концах многих хромосом Pan и Gorilla (как правило, на одних и тех же хромосомных плечах) выявляются небольшие Q-сегменты. У Pongo так же, как и у Homo, они отсутствуют. Существуют два возможных объяснения данного факта: либо эти сегменты - результат мутации de novo y общего предка Pan и Gorilla, либо они имелись у примитивного предка современных гоминоидов и были утра- чены в ходе эволюции Pongo и Homo. Обе гипотезы кажутся маловероятными, если исходить из классических представлений о дупликациях или делециях отдельных хромосомных сегментов как изолированных и случайных событиях; б) гетерохроматиновые районы выявляются в коротком плече некоторых акроцентрических хромосом. Их число уменьшается в следующем ряду организмов в последовательности: Pongo ® Gorilla ® Homo ® Pan. Повидимому, такой гетерохроматиновый материал время от времени образуется в виде мутаций de novo в непосредственной близости от центромер акроцентрических хромосом, а затем перемещается в результате случайных хромосомных перестроек в другие участки хромосом. Возможно, что подобный материал содержится во вторичной перетяжке 9-й хромосомы Homo (разд. 2.1.2). 9-е хромосомы человека и шимпанзе включают также гетерохроматиновый блок вблизи центромеры; в) существуют вариации и в локализации Т-сегментов. Единственное возможное объяснение этого факта состоит в том, что периодически материал Т-сегментов на концах хромосомы синтезируется de novo и затем распределяется по другим хромосомным участкам в результате перестроек; г) у горилл и человека выявлены дополнительные Q-сегменты вблизи центромер 3-й и 4-й хромосом; сходные сегменты обнаружены в хромосоме 9 гориллы, в хромосоме 13 человека и в 3-й хромосоме шимпанзе. У орангутана и гиббона их нет совсем. Все эти три результата, полученные при изучении терминальных Q- и Т-сегментов, гетерохроматиновых районов и прицентромерных Q-областей, свидетельствуют о том, что кариотипические различия между пятью рассматриваемыми близкородственными видами обусловлены не только перестройками хромосом, которые можно объ- Дата добавления: 2015-12-16 | Просмотры: 650 | Нарушение авторских прав |