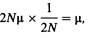|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Эволюция человека 21вистов. Отчасти они вызваны неправильным пониманием отдельных положений данной гипотезы, что убедительно разъясняется самим Кимурой [1941]. Чтобы понять эту гипотезу, необходимо уяснить два следующих момента. 1. В 1960-е годы была обнаружена громадная генетическая изменчивость на уровне белков и соответственно ДНК. С помощью методов определения аминокислотных последовательностей удалось выявить различия между гомологичными белками разных видов, а также между родственными белками одних и тех же видов. Изучение генетического кода вскрыло новые источники изменчивости, нуждающиеся в дальнейшем исследовании. Огромное количество ДНК, обнаруженное в эукариотической клетке (разд. 2.3.1.1), породило вопрос о функции избыточной ДНК и возможной причине этого феномена. Связаны ли большое количество ДНК и ее значительная изменчивость с естественным отбором, как это предполагалось неодарвиновской теорией эволюции, или же на молекулярном уровне большее значение имеют случайные процессы? Если бы решающим фактором был, как это предполагалось общепринятой синтетической теорией, отбор, то его действие испытывало бы огромное число сайтов ДНК. 2. Кимура разработал математические диффузионные модели, которые позволяют получить ответ на вопрос: «Какова вероятность того, что отдельный мутант, возникший в популяции конечного размера, рано или поздно распространится по всей популяции?» (т. е. оценить вероятность фиксации гена, разд. 6.4.1 и 6.4.2). В своей классической работе он сформулировал эту проблему и ее решение следующим образом: «Рассмотрим популяцию численностью в N особей.... Если мы заглянем достаточно далеко в будущее, то увидим, что вся популяция генов данного определенного локуса происходит от одного-единственного аллеля, имеющегося в нынешнем поколении. Это результат фатального процесса случайного генетического дрейфа1). Если в нынешнем поколении аллель А 1имеет частоту р, вероятность того, что удачливым аллелем, от которого произойдет вся популяция генов, будет A1, a не какой-нибудь другой аллель, равна ...р. Теперь, если мутация возникает с частотой μ на ген на поколение, то число новых мутантов по этому локусу в нынешнем поколении составляет 2Νμ2) Кроме того, вероятность, что данный ген рано или поздно зафиксируется в популяции, равна 1/2Ν. Таким образом, вероятность возникновения в этом поколении какого-либо мутантного гена и его фиксации в данной популяции равна
т. е. независимо от размера популяции частота нейтральной генной замены равна частоте мутаций». Позднее Кимура несколько скорректировал этот тезис, заметив, что мутант, селективно невыгодный в большой популяции, может быть нейтральным в небольшой и, таким образом, в малых популяциях скорость замен на самом деле будет выше. В своих модельных расчетах Кимура рассматривал случайное мутирование как процесс, независимый от времени. В широко известной монографии [1941] он пишет, что основываясь на положении о случайной фиксации мутаций, зависящей только от их частоты, можно сделать предсказание о линейности накопления генетических различий с течением времени, существующей независимо от изучаемого вида, продолжительности его поколения и других параметров. Пределы скоростей фиксации замен определялись только ограничениями, налагаемыми функциональными требованиями к генам и их продуктам-белкам: «отрицательный» отбор элиминировал макромолекулы, содержащие замены оснований и аминокислот, не совместимые с нормальным выполнением 1) Это положение следует из законов, лежащих в основе процесса случайной фиксации; разд. 6.4. 2) Где N = размер популяций. Дата добавления: 2015-12-16 | Просмотры: 576 | Нарушение авторских прав |