|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Нуклеиновые кислоты, строение и функции. Структурная модель ДНК Уотсона и Крика. Универсальность и видовая специфичность ДНК.Методы генетики. Гибридологический метод, его основные положения. Генетическая символика. Правила записи скрещивания.
Методы: 1)гибридологический- это основной метод, состоит в изучении наследования признаков. Эксперименты которые планируются позваляют проанализировать передачу признаков и св-в от родителей потомкам. Основные положения: скрещиваемые орг-мы должны принадлежать одному виду, орг-мы должны четко различаться по отдельным признакам, изучаемые признаки должны быть постоянны 2) цитогенетический, изучение кариотипа клеток орг-ма 3) малекулярно-генетический, начало этого метода положено в 1940-х годах на экспериментах бактериофага и бактерий. Были изучены репликация, регуляция и экспрессия (активность) генов, были клонированы гены 4) папуляционно-статистический, позволяет определить частоту встречаемости генов и генотипов в популяциях. Запись скрещивания: первая строка – Р (родители), гентип женского организма, знак скрещивания х, генотип мужского организма; вторая строка – буква G (гаметы) и (подобозначениями генотипов, в кружочках), гаметы женской и мужской особей; третья строка: буква Fk (потомки), генотипы потомков (под обозначениями генотипов могут быть указаны признаки организмов), k – номер поколения. Нуклеиновые кислоты, строение и функции. Структурная модель ДНК Уотсона и Крика. Универсальность и видовая специфичность ДНК.
Конечные продукты их гидролиза – пуриновые и пиримидиновые основания, пентозы и фосфорная кислота. По химическому составу различают дезоксирибонуклеиновою (ДНК) и рибонуклеиновую (РНК) кислоты. В состав ДНК входит моносахарид – дезоксирибоза, в состав РНК – рибоза. Эти соединения различаются между собой азотистыми основаниями, структурой молекул, клеточной локализацией, а также функциями. Соединения, молекула которых состоит из пуриновых или пиримидиновых оснований и пентозы (рибоза, дезоксирибоза), называются нуклозидами. Название нулеозида определяется азотистым соединением, которое входит в его структуру. Нуклеиновые кислоты строение и функции. Отдельные нуклеотиды соединяются между собой при этом образуют ди-, три-, тетра-, пента-, гекса, гепта- и полинуклеотиды, то есть НК. НК состоят из сотен и тысяч отдельных нуклеотидов, которые соединяются между собой при помощи гидроксильной группы, находящейся возле 3'-го атома Карбона пентозы одного нуклеотида с остатком фосфорной кислоты, который находится возле 5'-го атома Карбона пентозы следующего нуклеотида. ДНК являются основным генетическим материалом всех живых биосистем. В организмах, за исключением бактерий и вирусов, она локализируется в клеточных ядрах. Незначительное количество этой кислоты сконцентрировано в митохондриях и хлоропластах. РНК были идентифицированы практически в каждой клеточной фракции. Наибольшее количество РНК сосредоточено в рибонуклеопротеидных компонентах – рибосомах. Следует сказать, что основная масса РНК содержится в цитоплазме, и лишь 10–15 % входит в состав ядра. РНК с учетом клеточной локализации, биологической функции, молекулярной массы разделяют на три вида: рибосомальные, транспортные и матричные. рРНК локализируются в цитоплазматических гранулах рибосом, где они прочно связаны с белком. Они характеризируются высокой молекулярной массой. тРНК находятся в основном в гиалоплазме клеток, ядерной жидкости в митохондриях и хлоропластах. Они имеют небольшую молекулярную массу. Основной их функцией является транспортирование активированных аминокислот от комплекса аминокислота – АМФ-энзим к месту биосинтеза белка, то есть до рибосом. Научными исследованиями доказано, что каждая аминокислота имеет свою индивидуальную тРНК. На сегодня известно более 60 видов транспортных РНК. иРНК в процессе синтеза в ядре получает информацию с ДНК и переносит ее на рибосомы, где она реализуется при биосинтезе белка. Двойная спираль, структурная модель (гипотеза) дезоксирибонуклеиновой к-ты (ДНК), согласно к-рой молекула ДНК состоит из двух антипараллельных полинуклеотидных цепей, образующих правильную правозакрученную перевитую спираль и удерживаемых имеете водородными связями за счёт взаимодействия пар азотистых оснований. Предложена в 1953 Дж. Уотсоном и Ф. Криком. Свойство генетичекского кода:-универсальность – дискретность –специфичность(кодон кодирует только АК) –избыточность кода. 5. Природа генетического материала вироидов и вирусов. Механизмы вирусной инфекции. Особенности наследственных структур прокариот и эукариот. Спор о природе вирусов имеет не только теоретический интерес, но и большое практическое значение. Ясно, что мероприятия по борьбе с вирусными болезнями человека, животных и растений будут совершенно различными в зависимости от того, вызываются ли они живыми организмами, или же они возникают в результате попадания в организм или образования в нем каких-то веществ. Все мероприятия по борьбе с вирусными инфекциями исходят из признания живой природы вирусов. И практика доказала эффективность этих мероприятий. Вирусы являются возбудителями многих болезней. Респираторные инфекции вызывают более 200 вирусов, включая их серотипы, вирусы гриппа типов А, В, С. Распространенными вирусными инфекциями являются корь, герпетическая инфекция. В. полиомиелита продолжают циркулировать, но не вызывают паралитической формы заболевания благодаря плановой вакцинации. В определенных регионах распространены арбовирусные инфекции, передаваемые насекомыми (в основном клещами и комарами), в частности клещевой энцефалит. Возможны вирусные поражения разных органов и систем в результате постинфекционных процессов: миокардиты, панкреатиты, иммунодефицитные состояния.
6. Строение метафазных хромосом. Кариотип. Набор хромосом в соматических и половых клетках. Гомологичные хромосомы. Каждому из населяющих нашу планету видов растений и животных свойственно строгое число хромосом, обозначаемое 2n (диплоидный набор). В половых клетках число хромосом в два раза меньше и равно n (гаплоидный набор). В соматических клетках организма каждая хромосома имеет пара, идентичную как морфологически, так и генетически (гомологичные хромосомы). Исключение из этого правила составляют половые хромосомы у гетерогаметных особей. Специфический для определенного вида по числу и структуре набор хромосом получил название кариотипа. Хромосомы, определяющие пол особи, называют половыми хромосомами, а все остальные - аутосомами. Внутренне строение хромосом чрезвычайно сложно. По химическому составу они на 40% состоят из ДНК и на 60% из белков, в среднем около 60% из которых приходится гистоны. Строение метафазной хромосомы при исследовании с помощью светового микроскопа представляет следующим образом. Каждая хромосома состоит из двух хроматид, спирально закрученными и располагающихся параллельно оси хромосомы. Для прокрашивающихся в интерфазном яде участков хромосом используют термин "хромонема" - красящая нить. Утолщения на хромонемах получили название хромомер. В любой клетке имеется по две одинаковые хромосомы каждого сорта. Такие хромосомы получили название гомологичных (одинаковых) хромосом. Одну половину хромосом потомство получает от матери, а вторую (гомологичную) — от отца.
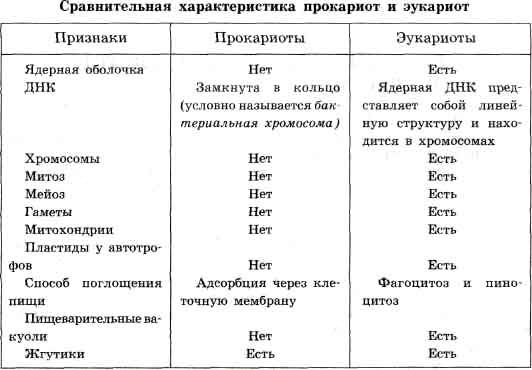
Деление клетки и воспроизведение. Механизм размножения прокариот. Распределение молекул ДНК при делении клетки прокариот. Митоз как механизм бесполого размножения эукариот. Генетическое значение митоза.
Постоянное деление клеток верхушечной и боковых меристем растений (камбия, пробкового камбия) обеспечивает длительный рост побегов и корней в длину и толщину. Вновь образованные клетки приобретают способность к делению после некоторого периода роста. Кроме того, делению предшествует удвоение клеточных структур: хлоропластов, митохондрий, центриолей и др. Перед началом клеточного деления должна реплицироваться ДНК, поскольку она несет в себе информацию, необходимую клетке для синтеза белков. Если бы дочерние клетки не получали точных копий ДНК материнской клетки, они перестали бы быть похожими друг на друга. Это в конечном итоге привело бы к невозможности существования определенных видов организмов. Чтобы этого не случилось, ДНК должна идеально реплицироваться и каждая дочерняя клетка при клеточном делении должна получать ее копию. Совокупность последовательных и взаимосвязанных процессов в период подготовки клетки к делению и в период деления называется митотическим циклом. Жизнь клетки от одного деления до следующего или до смерти называется клеточным или жизненным циклом клетки. У одноклеточных организмов клеточный цикл совпадает с жизнью особи. В непрерывно размножающихся тканевых клетках клеточный цикл совпадает с митотическим циклом и состоит из четырех периодов со строгой последовательностью смен друг друга: постмитотического (G1), синтетического (S), премитотического (G2) амитоза (М). Первые три периода —это интерфаза. По продолжительности она составляет большую часть митотического цикла клетки. В периоде G, клетка растет, в ней усиленно образуются РНК и белки, в первую очередь ферменты, катализирующие образование предшественников ДНК, ферменты метаболизма РНК и строительных белков. В 5-периоде происходит репликация ДНК (синтез новых цепочек ДНК — отсюда и название периода). В результате удвоения молекул ДНК в каждой хромосоме оказывается вдвое больше ДНК, чем было до S-периода, т. е. количество ДНК в диплоидных клетках соответствует тетраплоидному набору. Однако количество хромосом считается диплоидным, потому что образовавшиеся дочерние хромосомы (хроматиды) тесно переплетены между собой и составляют единую хромосому.
В S-периоде, кроме репликации ДНК, продолжается синтез РНК и белков, осуществляется синтез рРНК, которые используются в следующем периоде для синтеза белков, обеспечивающих митоз, происходит удвоение центриолей, а также деление митохондрий, увеличивается количество других органелл клетки. Время от окончания синтеза ДНК и до начала митоза называется G2-периодом. В этот период завершается подготовка клетки к митозу, активно синтезируются различные белки, в том числе и тубулиновые, из которых образуются нити веретена деления, а также иРНК и белки для осуществления (G1-периода после митоза). Далее наступает деление клетки. Деление клетки. После того как в клетке завершаются биохимические процессы подготовки к делению, начинается таинственный и до конца не изученный процесс. В настоящее время известно несколько способов деления клетки: митоз, прямое бинарное деление, амитоз и мейоз. Митоз (греч. mitos — нить), или непрямое деление, — основной способ деления эукариотических клеток. Митоз — это деление клетки, приводящее к образованию двух дочерних, в каждой из которых имеется точно такой же (аналогичный) набор хромосом, как и в родительской. Митоз — процесс непрерывный, но для удобства изучения биологи делят его на четыре стадии в зависимости от того, как выглядят в это время хромосомы в световом микроскопе. В митозе выделяют профазу, метафазу, анафазу и телофазу. В профазе происходит укорочение и утолщение хромосом вследствие их спирализации. В это время хромосомы двойные; они состоят из двух хроматид, связанных между собой в области первичной перетяжки кинетохором. Удвоение хромосом, как говорилось, произошло в 5-периоде интерфазы. Одновременно с утолщением хромосом исчезает ядрышко и фрагментируется (распадается на отдельные цистерны) ядерная оболочка. После этого хромосомы лежат в цитоплазме свободно и беспорядочно. В профазе центриоли расходятся к полюсам клетки, В конце профазы начинает образовываться веретено деления, которое формируется из микротрубочек. В метафазе завершается образование веретена деления, которое состоит из микротрубочек двух типов: хромосомных, которые с двух полюсов связываются с центромерами хромосом, и центросомных (полюсных), которые тянутся от полюса к полюсу клетки. Хромосомы как бы выталкиваются микротрубочками в область экватора клетки. В метафазе отчетливо видно двойное строение хромосом, соединенных только в области центромеры. В этот период легко подсчитывать число хромосом, изучать их особенности морфологического строения. В анафазе дочерние хромосомы (хроматиды) с помощью микротрубочек веретена деления расходятся и движутся к полюсам клетки. Во время движения дочерние хромосомы несколько изгибаются наподобие шпильки, концы которой повернуты в сторону экватора клетки. Таким образом, в анафазе хроматиды удвоенных в интерфазе хромосом расходятся к полюсам клетки. В этот момент в клетке находятся два диплоидных набора хромосом. В телофазе происходят процессы, обратные тем, которые наблюдаются в профазе: начинается деспирализация (раскручивание) хромосом, они набухают и становятся плохо различимы в световом микроскопе, Вокруг хромосом у каждого полюса из мембранных структур цитоплазмы формируется ядерная оболочка, в ядрах возникают ядрышки. Разрушается веретено деления. На стадии телофазы происходит разделение цитоплазмы с образованием двух клеток. В результате митоза из одной клетки возникают две дочерние с тем же набором хромосом. Таким образом, биологический смысл митоза состоит в строго одинаковом распределении между дочерними клетками материальных носителей наследственности — молекул ДНК, входящих в состав хромосом материнской клетки. Благодаря равномерному разделению реплицированных хромосом между дочерними клетками обеспечивается образование генетически равноценных клеток и сохраняется преемственность в ряду клеточных поколений. Это обеспечивает такие важные моменты жизнедеятельности, как эмбриональное развитие и рост организмов, восстановление органов и тканей после повреждения. Митотическое деление клеток является цитологической основой бесполого размножения организмов. Бактериальные клетки содержат только одну кольцевую молекулу ДНК, прикрепленную к клеточной мембране. Перед делением клетки ДНК реплицируется и образуются две идентичные молекулы ДНК, каждая из которых также прикреплена к клеточной мембране. При делении клетки мембрана врастает между двумя молекулами ДНК так, что в конечном итоге в каждой дочерней клетке оказывается по одной идентичной молекуле ДНК. Такой процесс получил название прямого бинарного деления. Амитоз или прямое деление, — это деление интерфазного ядра путем перетяжки. При амитозе веретено деления не образуется и хромосомы в световом микроскопе неразличимы. Такое деление встречается у одноклеточных организмов (например, так делятся большие полиплоидные ядра инфузорий), а также в некоторых высокоспециализированных с ослабленной физиологической активностью, дегенерирующих, обреченных на гибель клетках растений и животных либо при различных патологических процессах, таких как злокачественный рост, воспаление и т. п. Амитоз можно наблюдать в тканях растущего клубня картофеля, эндосперме семян, стенках завязи пестика и паренхиме черешков листьев. У животных и человека такой тип деления характерен для клеток печени, хрящей, роговицы глаза. При амитозе часто наблюдается только деление ядра: в этом случае могут возникнуть двух- и многоядерные клетки. Если же за делением ядра следует деление цитоплазмы, то распределение клеточных компонентов, как и ДНК, осуществляется произвольно. Амитоз в отличие от митоза является самым экономичным способом деления, так как энергетические затраты при этом весьма незначительны.
8. Мейоз как цитологическая основа образования и развития половых клеток. Генетическое значение мейоза. Типы мейоза (гаметный, споровый, зиготный). Мейоз I начинается с профазы I. Это наиболее продолжительная фаза мейоза, которая, в свою очередь, подразделяются на стадии лептотена, зиготена, пахитена, диплотена и диакинез. На стадии лептотены в ядре появляются слабоспирализованные хромосомы. Постепенно они приобретают нитевидную форму. Зиготена начинается с постепенно попарного соединения (конъюгации, синапсиса) по длине параллельно уложенных гомологичных хромосом. Соединение попарно хромосомы образуют биваленты. В связи с тем, что перед началом мейоза произошла редупликация хромосом, каждый бивалент состоит из четырех хроматид. Функцию синапсиса выполняет синаптонемный комплекс (СК) - белковое образование, входящие в состав бивалента и имеющие вид трехслойной ленты, располагающейся между конъюгирующими хромосомами. СК формируются постепенно по принципу застежки - молнии на протяжении всей стадии зиготены. Образование бивалентов создает предпосылки для возможности обмена гомологичными участками между гомологичными хромосомами (кроссинговера), что представляет важное генетическое событие. В то же время продолжается процесс конденсации хромосом. Пахитена - это стадия, на которой СК сформирован по всей длине хромосом (стадия стабильного синапсиса). Она характеризуется продолжающимся утолщением хромосом в результате непрерывной конденсации хроматина. На этой стадии происходит обмен гомологичным участками хроматид (кроссинговер) и, как следствие, рекомбинация сцепленных генов. На следующей за пахитеной стадии, получившей название диплотены, продолжается конденсация хромосом, но при этом начинается процесс расхождение гомологичных хромосом, которые удерживаются в точках обмена участками, возникшими при кроссинговере. Они получили название хиазм. Диакинез - последняя стадия профазы I. Она характеризуется максимальной конденсации хромосом. Исчезает ядрышко, а биваленты располагаются по периферии ядра. При этом гомологичные хромосомы удерживаются в составе бивалентов благодаря хиазмам. Далее следует метафаза I. Ее началу соответствует распад оболочки ядра и формирование веретена деления. Биваленты располагаются в экваториальной плоскости. Анафаза I - стадия, на которой гомологичные хромосомы расходится к полюсам. В результате число хромосом во вновь образующейся клетке (n) будет в два раза меньше, чем в родительской (2n). В этом отличие анафаза I мейоза от анафазы митоза. Окончательное расхождение хромосом к полюсам свидетельствует о том, что началась телофаза I. За ней у ряда видов следует очень короткий интеркинез, во время которого синтез ДНК и репликация хромосом не происходят, и начинается второе деление мейоза (мейоз II). В этом случае хромосомы не деконденсируются. Однако у некоторых видов растений интерфаза между первым и вторым делением мейоза продолжается довольно долго. В этом случае хромосомы деконденсируются, образуя два вида ядра, разделенные клеточной перегородкой. Второе деление мейоза протекает довольно быстро по типу обычного митоза, но уже в клетках с гаплоидным числом хромосом. В тех случаях, когда интерфаза короткая, профаза II выпадает и второе деление начинается с метафазы II, во время которой происходит образование веретена деления и хромосомы располагаются в экваториальной плоскости. В анафазе II центромеры делятся и начинается расхождение хроматид к полюсам, которое заканчивается на стадии телофазы II. На этой стадии происходит полная деконденсация хроматина, образуются ядра и клеточные перегородки. В конечном итоге в результате мейоза образуется 4 клетки, каждая из которых содержит в ядре гаплоидное (n) число хромосом. При зиготном типе мейоза, характерном для аскомицетов, базимицетов, некоторых водорослей, споровиков и др., для которых в жизненном цикле преобладает гаплоидная фаза, две клетки — гаметы — сливаются, образуя зиготу с двойным (диплоидным) набором хромосом. В таком виде диплоидная зигота (покоящаяся спора) приступает к мейозу, дважды делится, в результате образуются четыре гаплоидные клетки, которые продолжают размножаться. Споровый тип мейоза встречается у высших растений, клетки которых имеют диплоидный набор хромосом. В данном случае в органах размножения растений образовавшиеся после мейоза гаплоидные клетки еще несколько раз делятся. Другой тип мейоза, гаметный, происходит во время созревания гамет — предшественников зрелых половых клеток. Он встречается у многоклеточных животных, среди некоторых низших растений. В случае гаметного мейоза при развитии организма происходит выделение клонов герминативных клеток, которые впоследствии будут дифференцироваться в половые клетки. И только клетки этих клонов будут при созревании подвергаться мейозу и превращаться в половые клетки. Дата добавления: 2015-12-16 | Просмотры: 2029 | Нарушение авторских прав |