|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
ГЕНЕТИЧЕСКАЯ УСТОЙЧИВОСТЬ И ВОСПРИИМЧИВОСТЬ К ВИРУСНЫМ ИНФЕКЦИЯМЛейкозы (лейкемия, белокровие) — опухолевые заболевания кроветворной ткани. Характеризуются главным образом системным размножением незрелых кроветворных клеток в различных органах и тканях. Межвидовые и межпородные различия. Лейкоз чаще встречается у крупного рогатого скота, чем у4 овец, лошадей и свиней. У лошадей чаще регистрируются злокачественные меланомы. Среди 34 пород скота страны наиболее восприимчивы к лейкозу животные красных (бурая латвийская, красная датская, красная литовская и др.) и черно-пёстрых пород (Нахамеон, 1989). У пород швицкого происхождения (костромская, лебедин-ская, бурая карпатская) лейкозы появляются редко. Среди северонемецких пород лейкоз-1*егистрировался в 8 раз чаще, чем у баварских. Предполагают, что красная горбатовская порода, красный мегрельский, якутский, суксунский и бушуевский скот относительно устойчивы к лейкозу (Дмитриев и др., 1986). Ин-фицированность скатя молочных пород значительно выше, чем мясных. Однако некоторые исследователи считают, что в одинаковых условиях среды между породами отсутствуют различия по заболеваемости лейкозом. Влияние производителей и линий. Во всех породах между производителями выявлены большие различия по заболеваемости потомства лейкозом. Так, частота пораженное™ дочерей одних быков равна 0—5 %, других — 20—50 % и выше. Сила влияния отцов на устойчивость дочерей к лейкозу равна 15 %, а в некоторых стадах доля влияния составляет 3—5 %. От скрещивания резистентных отцов со здоровыми и лейкозными матерями получено в 3 раза меньше больных дочерей, чем от восприимчивых к лейкозу быков (табл. 65). 65. Заболеваемость лейкозом дочерей резистентных и восприимчивых к лейкозу быков (по В. Л. Петухову) Коэффициент ранговой корреляции быков, использовавшихся в разных хозяйствах, изменялся от 0,40 до 0,75. От отцов, заболеваемость дочерей которых была в пределах от 0 до 10 %, получены сыновья, давшие потомство также с относительно низкой заболеваемостью (5,4 %). При высокой пораженное™ отцов (30 % и более) дочери сыновей также характеризовались повышенной восприимчивостью к лейкозу (27,5). В хозяйствах может быть 21—28 % быков с относительно низкой заболеваемостью (до 5—10 %) и 13—23 % быков с высокой пораженностью потомства (больше 30 %). В пределах отдельных хозяйств выявлены межлинейные различия по частоте заболеваемости лейкозом. Однако ранг линий в разных хозяйствах и районах сильно изменяется. Только 0,8— 2,6 % дисперсии от общей изменчивости признака обусловлено влиянием линии. У мышей выведены линии, резко различающиеся по заболеваемости лейкозом и другими злокачественными новообразованиями. Влияние семейств и матерей. Существуют большие различия между семействами по частоте заболеваемости лейкозом. Имеются семейства, свободные от лейкоза, и с заболеваемостью до 70 % (Эрнст и др., 1989). В некоторых популяциях выявлено до 30 % свободных от лейкоза семейств и до 13 % семейств с пораженностью свыше 40 %. На рисунке 66 приведена генеалогическая схема восприимчивого к лейкозу семейства. От больных лейкозом матерей получено в 2 раза больше больных дочерей, чем от здоровых (табл. 66). Заболеваемость внучек в меньшей степени зависит от состояния здоровья бабушек. Коэффициент корреляции частоты заболеваемости лейкозом матерей и дочерей равен 0,2. Между возрастом проявления лейкоза у матерей и дочерей существует положительная связь (г = 0,39). Некоторые отцы также различаются по возрасту проявления лейкоза у дочерей. Заболеваемость лейкозом до 3 лет ниже, чем пораженность в более старшем возрасте. 66. Чистота заболеваемости потомства лейкозом в зависимости от состояния здоровья родителей (по В. Л. Петухову)
Конкордантность однополых двоен по лейкозу равна 74 %. Более близкие родственники лейкозных близнецов болеют чаще, чем более далекие. А. Г. Незавитин (1992) в племенных стадах выявил, что от РИД (—) матерей получено в 2 раза меньше РИД (+) дочерей, чем от инфицированных ВЛКРС матерей (табл. 67). vnk ^n J£S?i2!Z?*** схем* "Речисиоломеиного к лейкозу семейспа Азартной Л7М (первые цифры, нмшсшаые через -nipe, обозначают год ровдешы, ггорые -год выбыли, шшрвмер 72-75) 67. Частота инфицированности ВЛКРС дочерей, полученных от инфицированных и здоровых матерей (по А. Г. Незавитину)
Связь с генетическими полиморфными системами. Доказана разная по величине ассоциация главного комплекса гистосовместимости с некоторыми болезнями у человека и животных. Показано, что у джерсейского скота антигены МНС W1 и W3 связаны с устойчивостью к лейкозу. По мере инфицирования животных вирусом лейкоза в группе положительного по гематологии скота увеличивается частота антигенов BoLA W8 и W11. У лейкозных коров чаще встречаются аллели групп крови BG2O3Y2, Z, а у здоровых — BG2O3, BO3PFS2. Предполагают, что восприимчивость к лейкозу связана с некоторыми антигенами генетических систем групп крови. Животные бурой латвийской породы с типом церулоплазмина ВВ имеют более низкую заболеваемость лейкозом (1,27 %), чем животные с другими типами (Карликов, 1976). Анализируя все имеющиеся данные по ассоциации полиморфных систем с лейкозом, можно сказать, что пока нет генетических маркеров устойчивости и восприимчивости скота к лейкозу. Цитогенетика лейкоза. Во многих исследованиях обнаружены изменения кариотипа при лейкозах. При всех формах лейкоза крупного рогатого скота часто выявляется повышенный процент анеуплоидии. Изучение кариотипа костного мозга у лейкозных и здоровых животных показало, что у больного скота в 4 раза больше полиплоидных клеток. Однако в некоторых работах отсутствуют различия по частоте хромосомных аномалий у больных и здоровых животных. Пока четко не доказано, являются ли хромосомные аномалии одной из причин лейкоза. Наследование устойчивости и восприимчивости к лейкозу. Показано, что моногенное наследование устойчивости и восприимчивости лейкоза маловероятно. Распределение семейств и быков по заболеваемости потомства лейкозом отличалось от нормального, тогда как в этих же стадах по туберкулезу и бруцеллезу было нормальным. В некоторых исследованиях установлен нормальный характер распределения быков по заболеваемости дочерей лейкозом. Можно предположить, что устойчивость к лейкозу определяется многими генными локусами. Коэффициенты наследуемости устойчивости и восприимчивости к лейкозу в разных стадах изменяются от 0,07 до 0,5 (Эрнст, Петухов, Карликов и др.). Предполагают, что у кур восприимчивость к заражению вирусом лейкоза доминирует над резистентностью и является моногенным признаком. Локусы, определяющие устойчивость против вирусов лейкоза подгрупп А и В, наследуются независимо друг от друга. Гипотезы и теории этиологии лейкозов. Постоянно расширяются знания о злокачественных новообразованиях. Л. А. Зильбер создал вирусогенетическую теорию возникновения опухолей, согласно которой нуклеиновая кислота вируса частично или полностью включается в. геном клетки. О. А. Иванова (1972) считала, что лейкоз обусловлен прови-русом (V), ДНК которого входит в геном неблагополучного животного. В геноме также имеется ген-репрессор (R), влияющий на активность провируса. Заболевают особи, у которых имеются провирус и неактивный рецессивный аллель репрессора Wrr, Vvrr и частично WRr. Лейкемогенез — многостадийный процесс. Считают, что лейкоз вызывается РНК-с эдержащим вирусом лейкоза крупного рогатого скота (ВЛКРС или WmV), который относится к группе ретровирусов. Этот вирус интегрирован с геномом кроветворной клетки и способен к горизонтальной и вертикальной передаче. В ретровирусах за индукцию рака ответствен онкоген. Онкогены ретровирусов имеют не вирусное, а клеточное происхождение. Предполагают, что онкоген (протоонкоген) — это измененный нормальный ген. При попадании в геном ретровируса протоонкоген активизируется и превращается в онкоген, который может трансформировать клетки. Известно 17 протоонкогенов. Однако у группы ретровирусов лимфоидного лейкоза крупного рогатого скота онкогены не обнаружены. В. П. Шишков (1976) выдвинул вирусоиммуногенетическую концепцию этиологии патогенеза и профилактики лейкозов и ряда опухолевых заболеваний. Согласно этой теории наряду с ведущей ролью РНК-содержащих вирусов в развитии лейкозов крупного рогатого скота и птицы имеют значение иммунологическое состояние организма и генетическая предрасположенность к этой болезни. Для диагностики лейкоза широко применяют реакцию имму-нодиффузии (РИД). Во многих стадах может быть большой процент инфицированных животных. Далеко не всегда инфицированные ВЛКРС животные заболевают. У этих животных сохраняется нормальный уровень продуктивности. Однако 90—100 % заболевших лейкозом животных инфицированы ВЛКРС. Селекция животных на устойчивость к лейкозу может проводиться только в стадах с большой инфицированностью скота вирусом лейкоза. Рак глаз. Эта болезнь крупного рогатого скота распространена в Австралии. Считают, что ультрафиолетовые лучи — главный фактор в возникновении болезни. Наиболее чувствительны к раку глаз породы герефорд, комолый герефорд и их помеси. Болезнь у животных встречается в возрасте 8—10 лет и очень редко — до 5 лет. У животных с полной пигментацией кожи (породы абердин-ангус, гернсейская, джерсейская, швицкая и др.) рак глаз не зарегистрирован. У животных с уменьшением пигментации радужной оболочки заболевание встречается чаще (табл. 68). 68. Рак глаз и пигментация радужной оболочки (по Nishimura и др.)
Среди 353 помесей герефорд х шароле у 13,1 % животных пигментация полностью отсутствует, поэтому у них более высокая частота рака глаз. Ящур. Острая вирусная чрезвычайно контагиозная болезнь парнокопытных. К болезни относительно устойчивы зебу. На одной ферме из 1074 голов крупного рогатого скота заболело ящуром 97,1 %. Частота заболеваемости животных симментальской и красной степной пород равна 100 %, кавказской бурой — 87,5, а зебу — только 20 %. Имеются данные о семейной устойчивости скота к болезни. Болезнь Марека (MD) — инфекционная болезнь птиц (возбудитель — ДНК-содержащйй вирус), характеризующаяся контаги-озностью, разрастанием лимфоретикулярной ткани во внутренних органах, коже, мышцах, поражением периферических нервных стволов. Некоторые породы кур различаются по устойчивости к болезни Марека. Выявлены также и межлинейные различия (табл. 69). Во многих породах созданы резистентные линии кур. Так, английскими учеными создана линия кур породы род-айланд со 100%-ной резистентностью к болезни. Резистентные и восприимчивые линии различаются и по продукции интерферона, выполняющего защитную функцию при вирусных инфекциях. 69. Резистентностъ к болезни Марека инбредных линий кур и их кроссоа после экспериментального заражения (по Gavora, Spenser)
По мнению некоторых ученых, резистентность к болезни доминирует над восприимчивостью. При этом основную роль играют 2—4 аутосомных гена трех локусов. Установлено неполное доминирование резистентности над восприимчивостью. Сцепленные с полом гены существенно не влияют на резистентность, хотя смертность от болезни среди женских особей выше, чем среди мужских. Имеется довольно четкая взаимосвязь аллеля В21 группы крови В с резистентностью к болезни Марека (табл. 70). Почти все особи резистентной линии N были гомозиготны по антигену В21, в то время как большинство особей из восприимчивой линии Р имели антиген В19. 70. Зависимость резистентности кур к болезни Мврека от внтигенв В21 (по Hutt)
Обозначение: В~ — другие В-аллели, но не В21. Существует точка зрения о наличии по меньшей мере двух форм генетической резистентное™. Одна из них связана с некоторыми аллелями В-системы групп крови, особенно В21, и выражается повышенной иммуногенетической реактивностью и возрастной устойчивостью. Вторая форма устойчивости связана с локусами Th-1 и Ly-4 и определяется резистентностью лимфоид-ных клеток против инфекции. Описано много хромосомных нарушений в клетках, инфицированных вирусами. При MD анеуплоидия и разрывы хромосом — наиболее часто наблюдаемые аномалии. Так, пропорция анеугоюидных метафаз в лимфоцитах была 10 %, а в клетках костного мозга— 12 % по сравнению с 1,7 % в контроле. Некоторые исследователи считают, что нет значительной связи между признаками продуктивности и резистентностью к MD. В других исследованиях обнаружена отрицательная корреляция между резистентностью к MD и живой массой птицы, массой яиц, но положительная с яйценоскостью. Ныокаслская болезнь (псевдочума птиц) — высококонтагиозная вирусная болезнь главным образом куриных. Характеризуется пневмонией, энцефалитом и множественными точечными геморрагическими поражениями внутренних органов. При остром течении смертность среди молодняка достигает 100 %. У кур обнаружены генетические различия по устойчивости и восприимчивости. В опытах Франкис (1955) после заражения смертность кур в семействе А была 30 %, а в семействе D — 78 %. Кроме того, смертность после заражения более быстро увеличивалась у особей семейства D. При иммунизации кур вакциной с инактивированным вирусом, вызывающим ньюкаелскую болезнь, выявлены высокодостоверные различия между семьями по отцу. Коэффициент наследуемости силы иммунного ответа был 0,41. Птицы, лишенные Т-лимфоцитов, очень восприимчивы к болезни. При агаммаглобулинемии цыплята также чувствительны к болезни. Японские перепелки использовались как модельный объект селекции на резистентность к ньюкаслской болезни. В результате пораженность перепелок в V—VI поколениях снизилась с 41,5 до 2,05 и 2,63 %. Реализованный коэффициент наследуемости равен 0,6. В селекционируемой на резистентность группе смертность снизилась в несколько раз, повысились выводимость и живая масса птицы. Скрепи (почесуха) — медленно развивающаяся инфекционная болезнь, ведущая к дистрофическим изменениям центральной нервной системы. Вызывается вирусоподобным агентом, природа которого не выяснена. Пока не найдены антигены против возбудителей медленных инфекций (скрепи, висна, куру и др.). Болеют овцы в возрасте 2,5—4,5 года. Характерный признак болезни — зуд. Овцы стирают шерсть об изгороди, и на теле появляются голые места. Болезнь встречается на всех континентах и во многих странах, особенно в Австралии, Новой Зеландии, Южной Африке и США В Западной Европе скрепи регистрируется в течение 200 лет. Овцы пород рамбулье, гемпшир и тарги более устойчивы к скрепи (соответственно 14, 18 и 19 %), чем новозеландские овцы породы суффольк (39 %) и шотландские черноголовые (28— 43 %). Козы молочных пород чаще заболевают скрепи (61 %), чем ангорские (26 %). Эффективен был отбор линий английских пород овец шевиот и хердвик на увеличение (позитивная линия) и уменьшение (негативная линия) заболеваемости скрепи в ответ на подкожное введение агента скрепи, обозначаемого как SSBP// (табл. 71). 71. Скрепи у овец, вызванная заражением вгентом SSBP// (цит. по R. Kimberlin)
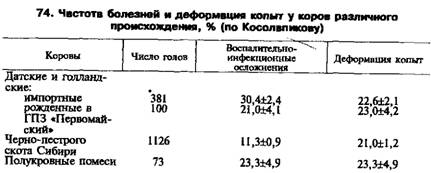
Инкубационный период у животных негативной линии во много раз больше, чем у животных позитивной. Предполагают, что ответ на заражение контролируется геном, обозначаемым sip (скрепи, инкубационный период) и имеющим две аллеля. Доминантный аллель контролирует восприимчивость к скрепи, а рецессивный — устойчивость. Следует учитывать тот факт, что ответ одной линии на разные агенты скрепи неодинаков. Например, заболеваемость овец породы шевиот позитивной и негативной линий на агент скрепи SSBP// была равна соответственно 100 и 50 %. Заболеваемость этих линий при внутрикожном заражении агентом скрепи СН 1641 была иной: соответственно 53 и 38 %. Все же заболеваемость животных негативной линии в обоих случаях была ниже, чем животных позитивной. Это говорит о возможности создания линий животных, устойчивых к нескольким штаммам одного возбудителя. Генетический контроль устойчивости к скрепи зависит окдггамма возбудителя. Миксоматоз кроликов. Острая вирусная болезнь, характеризующаяся серозно-гнойным конъюнктивитом и образованием опухолей в области головы, ануса и наружных половых органов. У некоторых пород кроликов смертность достигает 100 %. Возможность увеличения доли резистентности особей в популяции видна из следующего примера. В 1859 г. в Австралии были выпущены'! 3 кроликов, завезенных из Европы. В 1953 г. их насчитывалось около 0,5—1 млрд. Они стали приносить вред сельскому хозяйству. В 1950—1951 гг. в некоторых областях Австралии кроликов заразили вирусом миксоматоза, и через некоторое время погибло до 90 % особей. Однако проблема не была решена, так как среди кроликов возникли генетически резистентные особи и их поголовье сильно возросло. Успешна селекция на резистент-ность к миксоматозу и домашних пород кроликов. Коэффициент наследуемости устойчивости к этой болезни равен 0,35—0,40. Билет 47. Известно, что практически все реакции организма (нормальные и патологические) определяются индивидуальным генотипическим фоном. Генетические факторы, имея этиологическое значение, определяют развитие патологического процесса лишь тогда, когда действуют однонаправленно вместе с определенной совокупностью экзогенных (внешнесредовых) факторов. Язвенная болезнь является в целом мультифакторным заболеванием с полигенным типом наследования — это болезнь с наследственной предрасположенностью. С генетической точки зрения, язвенная болезнь представляет собой гетерогенную группу заболеваний, различающихся по степени генетической отягощенности, причем наиболее отягощенной является детская форма заболевания, а у взрослых наиболее тяжело протекает язвенная болезнь с локализацией в пилородуоденальной зоне. Обнаружение моногенных язвенных синдромов свидетельствует о том, что понятие “генетическая гетерогенная язвенная болезнь” включает не только различные по степени генетической отягощенности формы заболевания, но и формы с различным типом наследования, хотя надо полагать, что число подобных моногенных форм язвенной болезни, вероятно, незначительно, и в основной массе случаи язвенной болезни имеют полиэтиологическую природу, т.е. реализуются при совместном действии генетических факторов и факторов внешней среды. Наследственный компонент при язвенной болезни (особенно в случае пилородуоденальной локализации язвы) связан, вероятно, со значительным числом генов, каждый из которых определяет сам по себе наличие “нормального” признака, а в совокупности с другими такими же признаками способствует достижению некоторого “порогового” уровня — наличию генетической подверженности язвенной болезни. Выявленные достоверные ассоциации заболевания с менделирующими признаками заставляют рассматривать эти признаки в качестве факторов риска развития язвенной болезни. К доказанным генетическим факторам относят показатели максимальной секреции соляной кислоты; содержание пепсиногена I в сыворотке крови; повышение высвобождения гастрина в ответ на принятие пищи. К числу факторов, предположительно имеющих наследственную основу в развитии заболевания, относят следующие: 1. содержание пепсиногена II в сыворотке крови; 2. увеличение выброса соляной кислоты после еды; 3. повышение чувствительности обкладочных клеток к гастрину; 4. нарушение механизма обратной связи между выработкой соляной кислоты и освобождением гастрина; 5. расстройства моторной функции желудка и двенадцатиперстной кишки; 6. снижение активности фермента Z1-антитрипсина; 7. содержание адреналина в плазме и ацетилхолинэстеразы в сыворотке крови и эритроцитах; 8. нарушение выработки иммуноглобулина А; 9. морфологические изменения слизистой оболочки (гастрит, дуоденит) и некоторые другие. Генетическими маркерами развития язвенной болезни желудка и двенадцатиперстной кишки являются некоторые группы крови и особенности фенотипа. Существенное место в ряду генетических факторов язвенной болезни отводится способности секретировать антигены системе АВН, выявлению HZA-антигена, гистосовместимости В5, В15, В35. В настоящее время на базе наследственно-конституциональных факторов сформулирована гетерогенная теория, согласно которой в основе ульцерогенеза лежит полиморфизм пептических язв, сочетания различных генетически предопределенных факторов, обусловливающих полиморфизм клинических проявлений язвенной болезни. Билет 48. В условиях промышленного животноводства заболевания конечностей — наиболее распространенные болезни. От 3 до 7 % потерь молока связано с болезнями копыт у коров. В некоторых странах выбраковка из-за болезней конечностей составляет 2,4— 15,0 %. У голландского скота дефектам копыт, обусловливающим хромоту, подвержены 6—7 % молочных коров. Животные чаще заболевают в возрасте 4—7 лет. Десятки болезней конечностей часто объединяют под общим названием «болезни конечностей». Условия среды играют главную роль в их возникновении, однако наследственность — часто важный предрасполагающий фактор. Описан ряд моногенных наследственных болезней конечностей, но болезни с наследственным предрасположением занимают очень большое место. Серая местная и болгарская породы скота отличаются крепкой конституцией и особенно крепкими ногами и копытами. Заболеваемость коров черно-пестрой эстонской породы в 2 раза выше, чем красной эстонской. Изучена крепость копыт черно-пестрого скота и его помесей с голштинами. Помеси шведских быков характеризовались наибольшей крепостью копыт. Наименьшая крепость копыт была у помесей голштинских быков из США, Новой Зеландии и Голландии. Черно-пестрый скот Сибири менее подвержен инфекционным осложнениям отдела конечностей (11,3 %), чем импортный датский и голландский скот (21, 30,4 %) и их полукровные помеси (табл. 74).
Выявлены достоверные различия в частоте заболеваемости конечностей дочерей быков черно-пестрой породы. Так, в одном из стад пораженность потомства отдельных отцов изменялась от 0 до 23 % при средней заболеваемости по стаду 9 %. Коэффициент наследуемости общей крепости конечностей равен 0,13. Меньше предрасположен к болезни скот черно-пестрой (молочно-мясной тип) и герефордской пород. Генетическое разнообразие по устойчивости к болезням конечностей разных форм неодинаково. Например, h2 паралича задних конечностей равен 0,3, а опухоли межкопытной щели (лимакс) — 0,2. Частота лимакса у дочерей разных быков была в пределах 11—53 % при средней по стаду 23 %. Различия между линиями недостоверны. Не обнаружено 'достоверных различий по частоте деформаций и воспалительно-инфекционных осложнений между инбредными и аутбредными животными. У коров с антигеном групп крови J\ обнаружено меньше воспалительно-инфекционных осложнений (4,4 %), чем у животных с антигеном Оо (19,5 96). 2 Определенную роль в скорости роста, устойчивости к стиранию, количестве роговых трубочек на единице площади копыт, влагопоглощающей способности копытного рога и т. д. играет наследственность. Так, наследуемость количества роговых трубочек равна 0,38—0,75, диаметра трубочек — 0,63, степени стирания копыт — 0,79, влагоемкости — 0,82. Поражения копыт связаны с наследственной предрасположенностью к образованию винтообразного рога и язв подошвы. Постановка конечностей может также влиять на заболеваемость. Например, при саблевидной постановке наблюдаются язвы и нагноения, образование остроугольных или плоских копыт, при Х-образной постановке может возникать абаксиальный гнойный ламинит. Спастический (синдром) парез (поражение тазовых конечностей). Болезнь характеризуется рецессивным поли генным наследованием с неполной пенетрантностью рецессивного фактора. В большинстве случаев рецессивный фактор передается через быков. У 10—30 быков станций искусственного осеменения может встречаться спастический синдром Особенно высокая частота болезни у голштинских и абердин-ангусских быков. В связи с широким использованием в нашей стране голштинских быков необходимо учитывать в племенной работе ранние (3,5— 6 мес) и поздние (3—8 лет) формы спастического синдрома и других болезней. В США 2,6 % из 12 387 быков пяти молочных и трех мясных пород были выбракованы из-за спастического синдрома. При этом частота заболевания была выше у быков айрширской и голштинской пород (соответственно 3,9 и 3,5 %). В некоторых странах 5—10 % быков были выбракованы из-за этой болезни. Существует мнение, что использование инбридинга увеличивает заболеваемость до 12—17 %. В трех поколениях быков венгерского пестрого скота линии Султана клиническая форма спастического пареза зарегистрирована у 7 из 10 производителей. Изучено 1028 потомков пята быков линии Султана в трех поколениях. Частота заболеваемости потомства клинически больных быков в Fj равна 43,5 %, в F2 — 31,7 и F3 — 31,3 %. Заболеваемость потомства клинически здоровых быков была в несколько раз ниже: в Fi — 29,1 %, в F2 — 14 и в F3 — 6,6 %. Заболеваемость спастическим парезом в некоторых семействах достигала 60 %. При стойловом содержании спастический парез поздней формы (3—8 лет) наблюдался у 58 % животных, а при пастбищном — у 42 % (Хамори, 1983). У свиней изучено 13 признаков, вызывающих слабость ног. Коэффициенты наследуемости этих признаков были от 0 до 0,56, а в среднем h2 слабости ног равен 0,07. Рентгенографические исследования беконных свиней позволили установить, что h2 остеохондрита конечностей равен 0,4. Существуют различия межпометные и между потомством разных хряков по частоте слабости ног и повреждения суставов. Разные породы овец отличаются по восприимчивости к копытной гнили, вызываемой Fusiformis nodosus. Частота заболеваемости чистопородных коимбаторов в Индии была 12,8 %, а корриделей — 61,1 %. У гибридов Fi и 3/4-кровных по корриделям копытная гниль зарегистрирована соответственно у 21,1 и 26,1 % овец. Во всех группах заболеваемость взрослых овец была в несколько раз выше, чем у ягнят. Передние конечности поражаются чаще, чем задние. Можно предположить, что устойчивость к копытной гнили неполно доминирует над восприимчивостью. Британские породы овец и их кроссы более резистентны к болезни, чем мериносы. Изучение распространения врожденной косолатюсти у свиней пород ландрас, йоркширская и дюрок показало, что у ландрасских хряков частота случаев врожденной косолапости была выше, чем у животных других пород. Частота врожденной косолапости среди потомства разных хряков колебалась от 0 до 22 %. При этом частота косолапости у хряков в 1,7 раза выше, чем у свинок. Масса тела у поросят с косолапостью при рождении была ниже, чем у здоровых. Сделан вывод о полигенной обусловленности врожденной косолапости. Организация дифференцированной диагностики болезней конечностей, оценка потомства производителей и семейств по устойчивости к этим болезням — необходимый элемент племенной работы. Билет 49. Бесплодие — нарушение воспроизводства потомства. Полное или частичное бесплодие зарегистрировано в Швеции у 65 % быков. В США и Канаде из 8887 быков молочных пород выбраковано 36 % из-за различных нарушений воспроизводительной способности. Различные формы бесплодия встречаются у 20—40 % коров. В 32 стадах коров голштинской породы в США метриты зарегистрированы у 18,2 % животных, кисты яичников —у 10,4, задержка последа —у 8,6, аборты — у 1,4 % животных. У норвежского красного скота фено-типические корреляции задержки последа, метритов, молочной лихорадки, маститов, кетоза с бесплодием соответственно равны 0,77; 0,73; 0,69; 0,68; 0,57. Это значит, что все указанные болезни в значительной степени влияют на возникновение бесплодия. Степень проявления воспроизводительной способности изменяется от нормальной плодовитости до абсолютного бесплодия. Нарушение воспроизводительной способности обусловлено многочисленными факторами среды и наследственностью. Однако невозможно четко разделить ненаследственные и наследственные нарушения плодовитости. Коэффициенты наследуемости бесплодия очень малы — от 0 до 0,1 (Завертяев, 1986). Однако различия в частоте-бесплодия потомства разных производителей колеблются от 0 до 30 %. Воспроизводительные качества у голштинских коров несколько хуже, чем у черно-пестрых. Голштинским коровам на один отел требуется 1,91 осеменения, черно-пестрым — 1,55 и красно-пестрым — 1,59. У белых телок (болезнь белых телок) шортгорнской породы и родственных им пород часто встречается бесплодие. У белых телок матка и влагалище недоразвиты, тогда как яичники и наружные половые органы развиты нормально. Такая форма бесплодия встречается у 10 % белых шортгорнских телок. В одном стаде шортгорнов частота бесплодия у красных, чалых и белых телок соответственно равна 1,1; 3,5 и 39,1 %.
Средневерхнебельгийский скот белой и голубой масти произошел от шортгорнов. Процент бесплодных телок у белого скота (14,9) в 3 раза выше, чем у голубого (4,2). Установлено также, что среди телок белой масти бесплодных насчитывается 10—15 %, среди телок чалой масти — 1—1,75, а среди черных телок — только 0,3 %. Гипоплазия (недоразвитие) яичников и семенников раньше часто наблюдалась у комолого скота Северной Швеции (около 17 %). Телки с двусторонней гипоплазией не приходят в охоту, а у быков с двусторонней гипоплазией гонад не образуется сперма. У шведского фьелльского скота найдена связь между частотой гипоплазии и распространенностью белой окраски по поверхности тела. У животных с распространенностью белой окраски на 0,9 части поверхности тела частота гипоплазии гонад была 13,5 %, а у животных с пигментацией больше 50 % поверхности тела заболевания не было. Следовательно, качественный признак в какой-то степени может быть маркером некоторых форм бесплодия. Из дефектов органов половой системы следует указать на крипторхизм, при котором один или два семенника лежат в брюшной полости, и мошоночную грыжу, при которой из-за большого диаметра пахового кольца, соединяющего брюшную полость с мошонкой, петли кишечника заходят в мошонку. Крипторхизм у крупного рогатого скота встречается реже, чем у лошадей, коз, свиней и овец. У жеребцов с односторонним крипторхизмом ошюдотворяемость кобыл была 23 %, а у нормальных — 53 %. Крипторхизм наиболее часто встречается у собак пород с укороченным черепом (у боксеров до 23 %). У овец тонкорунной породы прекос доля крипторхов (больше двусторонняя) среди баранов достигает 25 %. У свиней крипторхизм встречается у 0,2—0,4 % особей, а в некоторых популяциях — до 3-5%. Крипторхизм может быстро распространяться в популяциях овец. На некоторых фермах в Венгрии частота крипторхов достигла 25—30 %. Среди комолых баранов крипторхизм регистрировался у 75,6 % особей, а среди рогатых — 2,4 %. Гипоплазия семенников отмечена у 9 баранов из 141. Среди комолых животных гипоплазия была у 77,7 %, среди рогатых — у 11,5 %. У коз и свиней крипторхизм наследуется по аутосомно-рецессивному типу. Исследования, выполненные в США на свиньях пород польско-китайской и ландрас, показали, что коэффициент наследуемости мошоночной грыжи равен 0,15. Доля материнского влияния примерно равна такой же величине. Крипторхизм и мошоночная грыжа — признаки, ограниченные полом. Гермафродитизм (совмещение мужского и женского пола в одном организме) чаще встречается у коз и свиней и реже — у крупного рогатого скота, лошадей и овец. У всех гермафродитов обнаружены половые хромосомы XX. У одних особей имеется только ткань семенника, у других — ткани семенника и яичника, но_ половые клетки не образуются. Частота гермафродитов неодинакова у разных линий и пород. У белой длинноухой свиньи она равна 0,1 %. Интерсексы чаще встречаются у комолых коз. В одном из стад зааненских и тоггенбургских коз выявлено соответственно 11 и 6 % интерсексов. Интерсексами генетически были самки. В Швеции у свиней выявлено 0,21 % интерсексов. В США в одном исследовании зарегистрировано 0,53 % интерсексов. У ландрасов частота таких свиней составляет 1,5 %, а у белой беконной породы в Венгрии — 3,8 %. Интерсексуальные животные подлежат выбраковке. Частота мертворожденности и абортов в некоторых стадах может изменяться в больших пределах (соответственно от 1 до 10 % и от 0,5 до 5 % и более) и обусловлена многими генетическими и негенетическими факторами. Существуют иммунные явления во всех звеньях нормального воспроизведения в организмах самок и самцов. Одной из причин прена-тальных потерь может быть врожденное или приобретенное им-мунодефицитное состояние самок. Совпадение некоторых антигенов главного комплекса гистосовместимости может вызвать нарушение эмбрионального развития. У свиней в 8,5 % случаев наблюдается гибель эмбрионов в первые 15—20 дней после оплодотворения. Существуют межпородные различия по частоте мертворожденных, абортов и трудных отелов (табл. 75, 76). У красного датского скота процент трудных отелов и мертворожденных значительно выше, чем у англерского. Однако у первотелок число трудных отелов и мертворожденных выше, чем у коров. Между трудными отелами и отходом телят генетическая корреляция равна 0,66. У голштинских первотелок в США задержка последа была у 1 % животных, а трудные отелы — у 45 %. У первотелок" айрширской породы задержка последа была во много раз выше (12—22 %), а количество трудных отелов ниже (8—18 %). Для снижения частоты затрудненных отелов необходимо использовать производителей с низким риском возникновения этого нарушения. 75. Частота мертворожденных телят и абортов у коров (по В. Л. Петухову)
76. Частота мертворожденных и трудных отелов у некоторых пород Скандинавских стран и ФРГ (цит. по Дехтяреву и др.)
Установлено, что частота абортов выше у дочерей, матери которых были предрасположены к абортам. Коэффициенты наследуемости предрасположенности к абортам и мертворождаемости были в пределах 0—0,08, трудных отелов — 0,04—0,30. Доказано достоверное влияние генотипов быков на частоту этих признаков. Пренатальная смертность у овец составляет 5—23 %. Большая смертность наблюдается у двоен. Один из методов борьбы с бесплодием — оценка генотипа производителей по этому показателю и широкое использование лучших из них. Дата добавления: 2015-12-16 | Просмотры: 5194 | Нарушение авторских прав |