|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
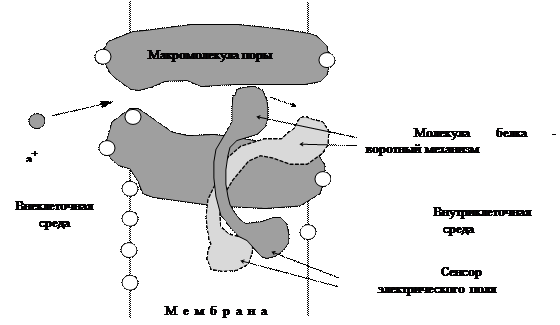
Характеристика канальных молекул.Распространение нервного импульса определяется присутствием в мембране нейрона энергетически управляемых Na+-каналов, открывание и закрывание которых ответственно за ПД. Хотя с химической точки зрения Na+-канал еще не достаточно изучен, известно, что он является белком с молекулярным весом от 25000 до 300000. Диаметр поры этого канала составляет 0,4-0,6 нм; через такую пору могут проходить ионы Na+, связанные с молекулами воды. На поверхности канала имеется много заряженных групп, размещенных в критических точках. Эти заряды обусловливают наличие большого электрического дипольного момента, который меняется по направлению и величине в соответствии с конформационными изменениями канала, сопровождающими переход из закрытого состояния в открытое. Поскольку поверхность мембраны клетки очень тонка, трансмембранная разность потенциалов в 70 мВ создает внутри мембраны сильное электрическое поле порядка 100 кВ/см. Подобно тому, как магнитные диполи имеют тенденцию ориентироваться вдоль силовых линий магнитного поля, электрические диполи белка натриевого канала стремятся встать параллельно линиям электрического поля мембраны. Изменения напряженности электрического поля могут переводить канал из закрытого состояния в открытое. По мере того, как под влиянием входящих ионов Na+ внутренняя поверхность мембраны становится более положительной, Na+-каналы все дольше находятся в открытом состоянии. Процесс открывания Na+-каналов под влиянием изменения потенциала мембраны называют активацией Na+-каналов. Этот процесс останавливается благодаря развитию другого процесса, названного Na+-инактивацией. Трансмембранная разность потенциалов, явившаяся причиной открывания Na+-каналов, затем переводит их в особое закрытое конформационное состояние, отличное от состояния, характерного для канала в покое. Второе закрытое состояние, вызванное состоянием инактивации, развивается медленнее, чем процесс активации, они остаются короткое время открытыми. В состоянии инактивации каналы пребывают несколько миллисекунд, а затем возвращаются в нормальное состояние покоя (рис. 16). Стенки Na+-канала несут отрицательный заряд, который отталкивает анионы и облегчает проход Na+. Вход в Na+-канал - селективный фильтр, проницаемый только для ионов Na+. Схематическая модель Na+-канала в мембране.
Рис. 16. Внутри Na+-канала находится воротный механизм, контролируемый потенциалом - заряженный сенсор в липидной фазе мембраны. При деполяризации, заряды смещаются, что вызывает изменение молекулярной конформации сенсора, что открывает проход через канал для ионов Na+. Пора открывается только на короткое время, поскольку через 1 мс наступает инактивация натриевых каналов. Не ясно, закрывается ли пора тем же воротным механизмом, или работает особая инактивационная пробка. Аналогично работает и Na+-насос. Электрически управляемые каналы - электрогенные насосы, есть еще химически управляемые каналы, например, синапсы. Дата добавления: 2015-12-15 | Просмотры: 707 | Нарушение авторских прав |