|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
СВОЙСТВА СЕРДЕЧНОЙ МЫШЦЫОсновными свойствами сердечной мышцы, определяющими непрерывное ритмическое сокращение сердца в течение всей жизни организма, являются автоматия, возбудимость, проводимость и сократимость. Автоматия. Под автоматией понимают способность сердечной мышцы ритмически возбуждаться и сокращаться без каких-иибо внешних по отношению к сердцу воздействий, т.е. без участия нервной системы и гуморальных факторов, доставляемых к сердцу кровью.
16 -3389 Доказательством автоматии сердца послужили следующие наблюдения и эксперименты. Изолированное сердце, т. е. выведенное из организма и помещенное в питательный раствор, продолжает самопроизвольно сокращаться. Даже разрезанное на кусочки, оно сокращается в том же ритме, что и у здорового животного. Если у животного денер-вировать сердце, т. е. перерезать все нервные стволы, подходящие к сердцу, оно продолжает сокращаться. На способности работать без воздействия внешних раздражителей основана пересадка сердца. Оживление остановившегося сердца достигается восстановлением спонтанной активности сердца, его автоматии. В чем причина такого уникального свойства сердца? У большинства беспозвоночных животных автоматия связана с нервными ганглиями, расположенными вблизи сердца, т. е. имеет ней-рогенную природу. У всех же позвоночных животных и у части беспозвоночных автоматия сердца обусловлена не нервными, а мышечными клетками, которые самопроизвольно деполяризуются после каждого потенциала действия. Эти клетки называются пейсмекерами, или «задающими сердечный ритм», или водителями сердечного ритма. Такая теория автоматии сердца называется миогенной. Способностью к автоматии обладают атипичные мышечные клетки, составляющие проводящую систему сердца. Ведущую роль в автоматии играет синусный узел. Он обладает наиболее высокой активностью по сравнению с другими участками проводящей системы, частота импульсации в нем наиболее высокая, и он задает определенную частоту сокращения сердца в состоянии физиологического покоя. Такой ритм обычно называют синусным ритмом, а синусный узел — водителем ритма сердца первого порядка. Если отделить лигатурой синусный узел от предсердий (опыт Станниуса), то обычно сердце останавливается. Однако через некоторое время оно снова начинает сокращаться, но в более редком ритме. Этот ритм «задает» следующий узел проводящей системы — атриовентрикулярный. Более редкие сокращения сердца обусловлены тем, что возбудимость атриовентрикулярного узла меньше, чем синусного. Этот узел называют водителем ритма сердца второго порядка. Если же и атриовентрикулярный узел перестает генерировать возбуждение, то водителем ритма сердца становится пучок Гиса, но его возбудимость еще меньше; пучок Гиса называют водителем ритма третьего порядка. В обычных условиях атриовентрикулярный узел и пучок Гиса только проводят возбуждение от синусного узла. Их собственная автоматия как бы подавлена главным пейсмекером, и только при развитии патологического процесса, прекращающего функцию синусного узла, свой ритм навязывают нижележащие узлы. Они являются латентными, или скрытыми, или потенциальными пейсмекерами. Какова природа автоматии? Методами электрофизиологии установлено, что потенциал действия (ПД) клеток проводящей системы отличается от других мышечных и нервных клеток. Во время расслабления сердца — диастолы — начинается медленно нарастающая деполяризация мембраны, которая затем переходит в фазу быстрой деполяризации (рис. 6.3, А). Фаза реполяризации в пейс-мекерах довольно продолжительная, в пейсмекерах синусного узла она имеет выраженное плато вместо пика потенциала. Сразу после возвращения мембранного потенциала к уровню потенциала покоя снова начинается медленная диастолическая деполяризация мембраны, и когда разность потенциалов между наружной и внутренней поверхностями мембраны уменьшается до определенного критического, или порогового уровня, внезапно возникает новый крутой сдвиг электрического заряда клетки, что свидетельствует о ее возбуждении.
Интервал между двумя ПД зависит от длительности медленной диастолической деполяризации, ее величины и порогового уровня сердечного ПД. Если скорость деполяризации уменыиает-
16*
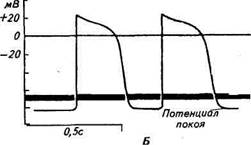
Медленная диастолическая деполяризация обусловлена особенностями ионной проницаемости мембраны пейсмекеров. Как и в других клетках, электрические процессы в мембранах миокарда являются следствием пассивного и активного перемещения ионов натрия и калия через тончайшие каналы (поры) в мембране, проницаемость которых регулируется заряженными частицами — ионами Са2+ или Мп2. Медленная диастолическая деполяризация объясняется тем, что во время реполяризации часть натриевых каналов не инактивируется и осуществляется медленный вход сначала натрия, а затем кальция в мембрану. Когда количество ионов натрия, проникших в клетку, снизит мембранный потенциал до критического уровня, наступает быстрая фаза деполяризации и ПД достигает своего максимального уровня. В теории об автоматии пейсмекеров еще много неясного, и раскрытие тончайших механизмов электрических процессов, происходящих в сердце, — актуальная задача современной кардиологии. Возбудимость. Возбудимость — свойство сердечной мышцы переходить в состояние возбуждения под влиянием различных раздражителей. В естественных условиях раздражителем является ПД, возникающий в синусном узле и распространяющийся по проводящей системе сердца до рабочих кардиомиоцитов. При некоторых заболеваниях сердца раздражение может возникать в других его участках, которые генерируют собственные ПД, и тогда сердечный ритм будет нарушен из-за взаимодействия разных по частоте и фазе ПД. В экспериментах на животных в качестве раздражителей могут быть использованы механические, термические или химические воздействия, если их величина превышает порог возбудимости сердца. При болезнях сердца, сопровождающихся нарушением сердечного ритма, больным вживляют в сердце миниатюрные электроды, питающиеся от батареек. Импульсы тока подаются непосредственно на сердце и возбуждают в нем ритмические импульсы. При внезапной остановке сердца или нарушении синхронизации отдельных мышечных волокон возможно воздействие на сердце прямо через кожный покров сильным коротким электрическим разрядом напряжением в несколько кВт. Это вызывает одновременное возбуждение всех мышечных волокон, после чего восстанавливается работа сердца. Во время возбуждения в сердце возникают физико-химические, морфологические и биохимические изменения, которые приводят к сокращению рабочего миокарда. Одними из ранних признаков возбуждения являются активация натриевых каналов и диффузия ионов натрия из межклеточной жидкости через мембрану, что приводит к ее деполяризации и возникновению ПД. В клетках рабочего миокарда ПД равен 80...90 мВ, при ПД Ю0...120мВ медленная диастолическая деполяризация в отличие от пейсмекеров отсутствует. Скорость нарастания деполяризации велика, восходящая часть ПД очень крутая, но реполяризация протекает замедленно, и мембрана остается деполяризованной в течение сотен миллисекунд (см. рис. 6.3, Б). Таким образом, длительность ПД в миокардиоцитах во много раз больше, чем в других мышечных волокнах. Благодаря этому все мышечные волокна предсердий или желудочков успевают сократиться до того, как какое-либо из этих волокон начнет расслабляться. Поэтому фаза реполяризации продолжается в течение всей систолы. Во время развития ПД возбудимость сердца, как и других возбудимых тканей, изменяется. Во время деполяризации возбудимость сердца резко снижается. Это — фаза абсолютной рефрактерности. Причиной ее является инактивация натриевых каналов, что прекращает поступление новых ионов натрия в мембрану. Если в скелетной мышце абсолютная рефрактерность очень кратковременная, измеряется десятыми долями миллисекунды и заканчивается в начале сокращения мышцы, то в сердце абсолютная невозбудимость продолжается весь период систолы. Практически это означает, что если во время систолы на сердце действует какой-либо раздражитель, даже сверхпороговый, то сердце на него не реагирует. Поэтому в отличие от скелетных мышц сердце не способно к тетаническим сокращениям и защищено от слишком быстрого повторного возбуждения и сокращения. Все сокращения сердечной мышцы одиночные. При очень большой частоте импульсов возбуждения сердце сокращается не на каждый ПД, а на только те из них, которые поступают по окончании абсолютной рефрактерности. Во время нисходящей фазы реполяризации, которая совпадает с началом расслабления сердечной мышцы, возбудимость сердца начинает восстанавливаться. Это — фаза относительной рефрактерности. Если в начале диастолы на сердце действует какой-либо дополнительный раздражитель, то сердце готово ответить на него новой волной возбуждения. Внеочередное возбуждение и сокращение сердца под действием раздражителя в период относительной рефрактерности называется экстрасистолой. Если очаг внеочередного возбуждения находится в синусном узле, то это приводит к преждевременному возникновению сер-
Рис. 6.4. Экстрасистола:
дечного цикла, при этом последовательность сокращений предсердий и желудочков не изменяется. Если же возбуждение возникает в желудочках, то после внеочередного сокращения (экстрасистолы) появляется удлиненная пауза. Интервал между экстрасистолой и следующей (очередной) систолой желудочков называется компенсаторной паузой (рис. 6.4.). Компенсаторная пауза объясняется тем, что экстрасистола, как и всякое сокращение сердечной мышцы, сопровождается рефрактерной паузой. Очередной импульс, возникающий в синусном узле, приходит в желудочки во время абсолютной рефрактерное™ и не вызывает их сокращения. Новое сокращение наступит лишь в ответ на следующий импульс, когда возбудимость миокарда восстановится. После относительной рефрактерности в сердце наступает очень короткий период повышенной возбудимости — экзальтации, когда сердце готово ответить даже на подпороговое раздражение. Проводимость. Проводимость — свойство сердечной мышцы проводить возбуждение. Как уже сказано, импульс возбуждения (ПД), возникая в пейс-мекерах синусного узла, распространяется сначала на предсердия. В предсердиях, где очень небольшое количество проводящих атипичных мышечных волокон, возбуждение распространяется не только по ним, но и по рабочим кардиомиоцитам. Это объясняет небольшую скорость распространения возбуждения в предсердиях. Поскольку синусный узел расположен в правом предсердии, а скорость передачи ПД невелика, то возбуждение правого предсер- дия начинается немного раньше, чем левого. Сокращение же левого и правого предсердий происходит одновременно. После того как возбуждение охватит мышцы предсердий, они сокращаются, а возбуждение концентрируется и задерживается в атриовентрикулярном узле. Атриовентрикулярная задержка длится до окончания сокращения предсердий, и только после этого возбуждение переходит на пучок Гиса. Таким образом, биологическое значение атриовентрикулярной задержки заключается в обеспечении последовательности сокращений предсердий и желудочков. Одновременное их сокращение иногда бывает при очень серьезной патологии, когда возбуждение возникает не в синусном узле, а в атриовентрикулярном и распространяется в обе стороны от атриовен-трикулярного узла — и в предсердия, и в желудочки. В таком случае наступает резкое нарушение гемодинамики в сердце. Механизмы атриовентрикулярной задержки не выяснены. Возможно, влияет низкая амплитуда ПД в клетках-пейсмекерах данного узла, сильная натриевая инактивация, большое сопротивление межклеточных контактов. Далее возбуждение распространяется по пучку Гиса, ножкам пучка Гиса и волокнам Пуркинье. Волокна Пуркинье контактируют с сократительными волокнами миокарда, и возбуждение передается с проводящей системы на рабочие мышцы. Скорость распространения возбуждения в сердце следующая: от синусного узла до атриовентрикулярного узла — 0,5...0,8 м/с; в атриовентрикулярном узле — 0,02...0,05; по проводящей системе желудочков — до 4,0; в сократительной мышце желудочков — 0,4 м/с. Непосредственная связь проводящей системы сердца с рабочими кардиомиоцитами осуществляется с помощью многочисленных разветвлений волокон Пуркинье. Передача сигналов происходит электрическим путем с небольшой задержкой. Эта задержка возбуждения способствует суммированию импульсов, неодновременно поступающих по волокнам Пуркинье, и обеспечивает лучшую синхронизацию процесса возбуждения рабочего миокарда. В рабочем миокарде имеются контакты как между торцами, так и боковыми поверхностями волокон. Поэтому возбуждение от основных стволов проводящей системы (ножек пучка Гиса) практически одновременно распространяется на правый и левый желудочки, обеспечивая их одновременное сокращение. Направление возбуждения внутри желудочков различно у животных разного вида. Так, у собак возбуждение вначале возникает на расстоянии нескольких миллиметров от внутренней поверхности мышечной стенки, а затем переходит к эндокарду и эпикарду. У копытных (у коз) направление распространения возбуждения в толще мышечной стенки меняется много раз, и множество волокон в районах эндокарда, эпикарда и в глубине стенки активируется практически одновременно.
В межжелудочковой перегородке возбуждение начинается в Особенности распространения возбуждения в сердце имеют значение при анализе электрокардиограммы — записи биотоков сердца. Сократимость. Сокращение — специфический признак возбуждения сердечной мышцы. Как и в других мышцах, сокращение сердечных мышечных волокон начинается после распространения потенциала действия по поверхности клеточных мембран и является функцией миофибрилл. Сократительная система миофиб-рилл представлена четырьмя белками — актином, миозином, тро-понином и тропомиозином. Сокращение миофибрилл сердца в принципе не отличается от сокращений скелетных мышц согласно теории скольжения протофибрилл Хаксли. Суть теории Хаксли заключается в скольжении тонких актино-вых нитей в промежутки между толстыми миозиновыми нитями,; что приводит к укорочению саркомера. При расслаблении мышцы актиновые нити отодвигаются назад, занимая исходное положение. В механизме скольжения актиновых нитей имеет значение кальций, депонированный в саркоплазматическом ретикулуме. Последовательность электрических и механических процессов при сокращении сердечных мышечных волокон в настоящее время представляется следующим образом. Потенциал действия, возникший на поверхности мембраны мышечного волокна, по поперечным Т-трубочкам, которые являются впячиваниями наружной мембраны, достигает системы поперечных трубочек, соединенных с цистернами саркоплазматического ретикулума. Полости сарко-плазматического ретикулума не сообщаются ни с Т-трубочками, ни с интерстициальной жидкостью и заполнены раствором с высоким содержанием ионов кальция. Полости Т-трубочек имеют такой же состав, что и межклеточная жидкость. Во время возбуждения активируются натриевые каналы в мембранах Т-трубочек и в миоплазму входят ионы натрия и кальция из межклеточной жидкости. Большая часть входящего кальция не участвует в сокращении миофибрилл, а пополняет его запасы в саркоплазматическом ретикулуме. Под воздействием потенциала действия повышается проницаемость мембраны саркоплазматического ретикулума и ионы кальция вьщеляются из него в миоплазму. Ионы кальция связываются с тропонином, что вызывает конформационные изменения в его молекуле. Сдвиг тропонин-тропомиозинового стержня I обеспечивает взаимодействие нитей актина и миозина (напомним, Щ что в расслабленной мышце актиновые волокна прикрыты молеку- 1 лами тропонина и тропомиозина, образующими комплекс, препятствующий скольжению протофибрилл). После освобождения актиновых нитей от блокировки тропо-миозиновым комплексом миозиновые головки присоединяются к соответствующему центру актиновых нитей под углом 90°. Затем наступает спонтанный поворот головки на 45°, развивается напряжение и происходит продвижение актиновой нити на один шаг. Эти процессы осуществляются за счет энергии АТФ, причем распад АТФ катализируется актомиозиновым комплексом, обладающим АТФ-азной активностью. Когда возбуждение прекращается, содержание ионов кальция в миоплазме снижается вследствие работы кальциевого насоса и закачивания кальция в саркоплазматический ретикулум, причем на работу кальциевого насоса также затрачивается энергия АТФ. В результате снижения содержания кальция в миоплазме тропо-миозиновый комплекс защищает активные центры актомиозино-вых нитей. Нити миозина и актина восстанавливают исходное положение, и мышца расслабляется. Изложенная теория сокращения сердечной мышцы во многом объясняет экспериментальные и клинические наблюдения о влиянии кальция и магния — его антагониста на работу сердца. Известно, что при перфузии изолированного сердца раствором, не содержащим кальция, оно останавливается, а при добавлении кальция в перфузионный раствор сокращения восстанавливаются. Известно также, что сердечные глюкозиды (например, препараты наперстянки) увеличивают проницаемость мембран для кальция и тем самым восстанавливают транспорт кальция между саркоплазматическим ретикулумом, наружной мембраной и миоплазмой. Согласуется с теорией мышечного сокращения и благоприятное влияние на сердце макроэргических веществ, энергия которых используется не только для механического сокращения, но и для работы ионных насосов — кальциевого и калиево-натриевого. Сократительные свойства сердечной мышцы несколько отличаются от скелетных. Если скелетная мышца реагирует на раздражение в соответствии с его силой, то сердечная мышца подчиняется закону Боудича «все или ничего». Его суть заключается в том, что на подпороговые раздражения сердце не сокращается («ничего»), а на пороговое раздражение отвечает максимальным сокращением («все»), и увеличение силы раздражителя не приводит к увеличению силы сокращения. В скелетных мышцах закону «все или ничего» подчиняются отдельные мышечные волокна. Дело в том, что потенциал действия вызывает освобождение кальция из саркоплазматического ретикулума равномерно по всей длине волокна, поэтому оно сокращается полностью. Но в скелетной мышце имеются волокна с разной степенью возбудимости, поэтому при слабом раздражении сокращаются не все волокна и суммарное сокращение оказывается небольшим. В сердечной же мышце волокна рабочего, т. е. сократительного, миокарда соединены межклеточными контактами (выростами плазматических мембран), что способствует практически одновременному распространению потенциала действия по всей мышце, и она возбуждается и сокращается как единый орган, 1 являясь функциональным синцитием. Закон Боудича является скорее правилом с определенными ограничениями. При подпороговом раздражении сокращение, действительно, не возникает, но в это время начинается активация натриевых каналов и повышается возбудимость миокардиоцитов. Возникающие местные потенциалы могут суммироваться и вызвать распространяющийся потенциал действия. С другой стороны, сила сокращения сердца, как хорошо известно, непостоянна и может изменяться в различных условиях жизни. Другая характерная особенность сердечной мышцы заключается в том, что сила сокращения сердца зависит от степени растяжения мышечных волокон во время диастолы, когда полости заполняются кровью. Это — закон Франка — Старлинга. Указанная закономерность объясняется тем, что при растяжении сердца кровью во время диастолы актиновые нити несколько вытягиваются из промежутков между миозиновыми, и при последующем сокращении возрастает число генерирующих силу поперечных мостиков. Кроме того, при растягивании сердечной мышцы в ней повышается сопротивление упругих элементов, и во время сокращения они играют роль «пружины», увеличивая силу сокращения. Особенно важное значение закон Франка — Старлинга имеет во время усиленной работы сердца, когда возрастает объем крови, поступающей в него во время диастолы. Увеличение силы сокращения приводит к тому, что вся кровь выбрасывается при систоле желудочков в артериальные сосуды, иначе после каждого сокращения в сердце оставалась бы значительная порция крови. При отсутствии большой нагрузки и небольшом объеме кровотока сила сокращения сердца умеренная. Таким образом сердце способно регулировать в известных пределах силу сокращения в зависимости от объема кровотока. Дата добавления: 2015-12-15 | Просмотры: 858 | Нарушение авторских прав |
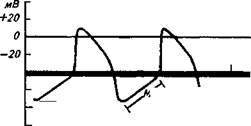
 ся (например, при охлаждении синусного узла), то пороговый уровень деполяризации наступает позднее, частота ПД и сокращений сердца уменьшаются. При возрастании скорости деполяризации мембраны, напротив, пороговый уровень деполяризации возникает раньше и это приводит к учащению возбуждения сердца. Отчасти этим объясняется учащение сердечной деятельности при повышении температуры тела.
ся (например, при охлаждении синусного узла), то пороговый уровень деполяризации наступает позднее, частота ПД и сокращений сердца уменьшаются. При возрастании скорости деполяризации мембраны, напротив, пороговый уровень деполяризации возникает раньше и это приводит к учащению возбуждения сердца. Отчасти этим объясняется учащение сердечной деятельности при повышении температуры тела.
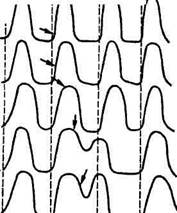 J, 2, 3 — искусственное раздражение наносят во время сокращения желудочков, поэтому эффекта нет (абсолютная рефрактерная фаза); 4, 5, б—раздражение наносят во время расслабления мышцы и вызывают появление экстрасистол, амплитуда которых тем больше, чем позже в период расслабления нанесено раздражение. Это связано с изменением возбудимости мышцы, наступающим после возбуждения; прямые стрелки и пунктирные линии указывают момент появления импульсов в синусном узле, боковые — момент искусственного раздражения сердца
J, 2, 3 — искусственное раздражение наносят во время сокращения желудочков, поэтому эффекта нет (абсолютная рефрактерная фаза); 4, 5, б—раздражение наносят во время расслабления мышцы и вызывают появление экстрасистол, амплитуда которых тем больше, чем позже в период расслабления нанесено раздражение. Это связано с изменением возбудимости мышцы, наступающим после возбуждения; прямые стрелки и пунктирные линии указывают момент появления импульсов в синусном узле, боковые — момент искусственного раздражения сердца