|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Айала Ф., Кайгер Дж. Современная генетика: В 3-х т. Т. 2. Пер. с англ.: – М.: Мир, 1988. – 368 с.172 Экспрессия генетического материала
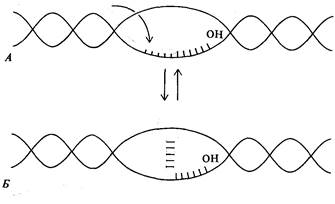
за счет смыкания участков матричной и комплементарной ей цепей ДНК. Кроме того, концевая последовательность РНК-транскрипта, состоящая в основном из остатков рибоуридина, и комплементарный полидезоксиаденозиновый участок матричной цепи ДНК образуют, как известно, не слишком прочную систему водородных связей. Это также может способствовать диссоциации РНК-транскрипта от матричной цепи ДНК.
Айала Ф., Кайгер Дж. Современная генетика: В 3-х т. Т. 2. Пер. с англ.: – М.: Мир, 1988. – 368 с. 15. Регуляция экспрессии генов у прокариот 173 Lac-оперон Генетический анализ системы утилизации лактозы клетками E.coli, осуществленный Франсуа Жакобом и Жаком Моно, можно назвать важнейшим шагом на пути к пониманию механизмов регуляции бактериальных генов. На базе этого исследования была впервые сформулирована модель структурно-функциональной организации оперона. Для утилизации лактозы клетке необходимы продукты двух тесно сцепленных цистронов. Это ген lacZ +, кодирующий ß-галактозидазу (рис. 15.1), и ген lacY+, кодирующий пермеазу, которая обеспечивает активный транспорт лактозы в клетку. Третий ген lac А +, кодирующий фермент тиогалактозидтрансацетилазу, регулируется координированно с генами lacZ+ и lacY +, но не имеет непосредственного отношения к утилизации лактозы. Эти гены картируются в последовательности Z-Y-A. Синхронная индукция всех трех белков реализуется через синтез одной общей полицистронной мРНК. Обычно в отсутствие индуктора уровень транскрипции генов lac очень невелик и в клетке имеются лишь очень малые количества ß-галактозидазы и пермеазы. В то же время удается легко отобрать мутанты (так называемые конститутивные мутанты), которые и в отсутствие индуктора продуцируют большие количества этих белков, такие же, как нормальные клетки в условиях индукции. Все эти конститутивные мутанты содержат мутации вблизи гена Z и на достаточном удалении от генов Y и А. Как показал комплементационный анализ, такие мутанты можно разбить на два класса: I – (неиндуцибельные) и 0c (операторно-конститутивные). С использованием элементов F'lac или конъюгативных мерозигот (см. гл. 8) можно получить частичные диплоиды, включающие такие мутации. Фенотипические проявления частичных диплоидов этого типа свидетельствуют, что мутации I – и 0с оказывают принципиально различное влияние на синтез ß-галактозидазы. Из перечисленных в таблице 15.1 частичных диплоидов два первых характеризуются нормальным индуцибельным фенотипом, что указывает на рецессивный характер мутации I –. Ген I + осуществляет нормальный контроль экспрессии гена Ζ + в случае обоих частично диплоидных генотипов независимо от того, находится ли ген Ζ + на той же хромосоме, что и I +, или на другой. В случае двух по-
Дата добавления: 2015-12-16 | Просмотры: 609 | Нарушение авторских прав |