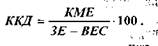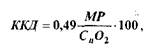|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Режими скорочення м’язових волокон.Міотонометрія В залежності від частоти імпульсації мотонейрона м’язові волокна рухової одиниці (РО) можуть працювати в двох режимах – в режимі поодинокого скорочення і в режимі тетанічного скорочення (мал.12). Режим поодиноких скорочень виникає тоді, коли інтервал між суміжними імпульсами мотонейрона дорівнює або дещо більшій тривалості поодинокого скорочення інервованих ним м’язових волокон. Робота м’язових волокон в режимі поодиноких скорочень відбувається при відносно низькій частоті імпульсації мотонейронів, яка не однакова у різних рухових одиниць. Це зумовлено перш за все різною тривалістю поодинокого скорочення різних м’язових волокон (мал. 13). Так, для повільних РО, м’язові волокна яких мають тривалість поодиноких скорочень 100 мс. (0,1 с), режим поодиноких скорочень спостерігається при частоті імпульсації мотонейрона, яка не перевищує 10 імп/с. Для швидких РО, м’язові волокна яких мають меншу тривалість поодинокого скорочення (наприклад, 20 мс), режим поодиноких скорочень спостерігається при частоті імпульсації мотонейрона – 50 імп/с. В кривій поодинокого ізометричного скорочення м’яза виділяють три фази: латентний (прихований) період збудження, фаза підйому напруження і фаза розслаблення. При ізотонічному скороченні ці фази позначаються дещо інакше: латентний період збудження, фаза вкорочення, фаза видовження. Тривалість фази напруження (вкорочення) приблизно в двічі коротша, ніж фаза розслаблення (видовження).
Мал. 13. Криві поодинокого скорочення змішаного м’яза (А), м’яза з білих (Б) і червоних (В) волокон: І – скорочення, II- струм дії.
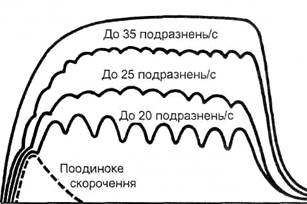
Тривалість фази вкорочення окремих РО, а, отже, швидкість скорочення різних м’язів у різних тварин не однакова. Так, для м’язів крила колібрі вона складає 8 мс, для згиначів пальців кішки – 10 мс, для скелетних м’язів людини – 10-70 мс, для литкового м’яза черепахи – 300 мс. В середньому час поодиноких скорочень РО різних м’язів ссавців коливається від 10 до 20 мс, а сила поодиноких скорочень – від десятої долі грама до 100 г. Працюючи в режимі поодиноких скорочень, м’язові волокна неспроможні розвинути велике напруження. Це пояснюється коротким періодом активного стану, внаслідок чого скоротливі елементи м’яза починають розслаблюватись, не досягнувши максимуму напруження. Тонічне напруження м’язів. Здатність скелетної мускулатури тривало, стійко підтримувати скорочення(напруження) називається тонусом, або тонічним напруженням. Робота м’язів в умовах тонічного напруження пов’язана з меншими енерговитратами, ніж при звичайному гладенькому тетанусі. Цим пояснюється низька втомлюваність м’язів, які знаходяться в тонусі. Тонус скелетних м’язів підтримується завдяки надходженню відносно нечастих імпульсів ЦНС (до 10 імпульсів за 1 с). Такі еферентні імпульси є проявом тонусу нервових центрів. Надходячи по мото-нейронах до м’язових волокон, вони викликають слабкий зубчастий тетанус, який і проявляється у вигляді енергоекономних тонічних скорочень м’язів. В режимі поодинокого скорочення працюють повільні РО, які включаються в активний стан по черзі, тривалий час підтримуючи позу тіла та протидіючи силам земного тяжіння. Тонус м’язів має рефлекторне походження. Його виникнення і підтримання зумовлене наявністю в м’язах тонічно напружених рухових одиниць, які постійно контролюються нервовими центрами. В свою чергу, тонус нервових центрів підтримується по зворотних зв’язках аферентними імпульсами, що йдуть від пропріорецепторів. М’язовий тонус є основою підтримання необхідної пози тіла в умовах постійної дії сил земного тяжіння. Дослідження м’язового тонусу є важливим елементом у вивченні стану опорно-рухомого апарату школярів, які займаються фізичною культурою та спортом. Особливо важливе значення має вивчення м’язового тонусу для нормування навантажень школярів. Розкриття регуляції тонуса скелетних м’язів у людини в умовах дії земного тяжіння, створює необхідні передумови для дослідження тонуса скелетних м’язів в умовах абсолютної і відносної невагомості. Адже майбутні польоти людей в космос вимагають знань того, як змінюватиметься координація рухів, в якій мірі та за яких умов вона зберігатиметься якнайдовше. Міотонометрія. Вивчення тонусу м’язів грунтується на вимірюванні напруження м’язів в даний момент часу. Звідси, тонус м’язів – це їх реакція на механічні впливи (дії) поздовжньої чи поперечної направленості. В залежності від цих впливів виділяють динамічний тонус, коли визначається опір м’язів, розтягнених в повздовжньому напрямку, і пластичний – коли визначається опір м’язу тиску, що діє в поперечному напрямку щодо певної точки м’язу. Амплітуда тонуса (АТ-різниця між показниками тонуса м’язу при його довільному скороченні і розслабленні) завжди більш висока у спортсменів, ніж у ненатренованих осіб. Чим вища амплітуда тонуса, тим вища натренованість спортсмена. AT різна у важкоатлетів та легкоатлетів, завжди нижча у осіб, які не займаються фізичною культурою і спортом. При перевтомі AT зменшується. Показник міотонометрії є об’єктивним методом оцінки функціонального стану м’язів спортсмена. Його зміни у різні періоди тренування доцільно використати для корекції тренувальних навантажень спортсменів. Існують вікові особливості тонусу скелетних м’язів. У хлопчиків 8-9-літнього віку він вищий, ніж у дівчаток. Суттєве збільшення тонусу скелетних м’язів відмічається у підлітків 12-15-літнього віку. (С.І.Гальперін, 1967). У ранньому віці скелетні м’язи знаходяться в стані постійного тонуса і рівень виходу актинових ниток за межі міозинових не суттєвий. В дошкільному віці, із зменшенням участі скелетних м’язів в теплоутворенні, вони набувають все більшої здатності до розслаблення. Як наслідок зростає рівень входження актинових ниток між міозинові, зростає показник амплітуди тонуса. Довільне розслаблення скелетних м’язів більш утруднене, ніж їх довільне напруження. Спроможність м’язів до розслаблення з віком зростає. Вимірювання тонуса м’язів в умовах максимального напруження і при розслабленні (міотонометрія) дозволяє кількісно характеризувати їх скоротливу функцію і здатність до розслаблення (еластичність, твердість, пружність). Скорочення та розслаблення окремих груп м’язів обумовлені досконалістю центральнонервових механізмів координації (див. "Регуляція напруження м’язів"). Міотонометричні дослідження згаданих процесів є важливим об’єктивним методом оперативної оцінки рівня функціонального стану нервово-м’язового апарата і впливу на нього різних за характером навантажень. Міотонометрію проводять з допомогою приладів Уфлянда, Жукова, Дубровського, Дерябіна, Козловського. Одиницею виміру тонуса м’язів є міотон (умовна одиниця шкали міотонометра). Міотони приладів різних конструкцій нерівноцінні, а тому для попередження помилок при порівняльній міотонометрії, необхідно користуватися одним типом приладів. Свідоме напруження чи розслаблення м’язів пов’язане з формуванням складних і тонких диференційовок. Ось чому у ненатренованих до фізичної роботи осіб спроможність гальмувати м’язовий тонус розвинута недостатньо (показник додаткового розслаблення складає 0,7-0,9), що свідчить про низьку здатність м’язів до розслаблення. Свідоме бажання знизити м’язовий тонус малоефективне. Відомо чимало випадків коли спроба учня розслаблювати м’язи, навпаки призводить до їх напруження. Кваліфіковані спортсмени володіють більшою здатністю як до напружень, так і до розслаблень м’язів (Ю.Г.Козловський, 1976, 1980). Здатність до активного розслаблення м’язів у спортсменів різних спеціалізацій різна. Найбільш низькі величини тонуса м’язів зустрічаються у легкоатлетів, гімнастів і єдиноборців, більш високі – у футболістів. Боксери добре володіють прямим м’язом живота. Тетанічне скорочення м’язів. При руховій діяльності звичайно до м’язових волокон надходить серія наступних нервових імпульсів, які викликають тривале тетанічне скорочення м’язів – тетанус. Тетанічні скорочення властиві тільки скелетним м’язам і не характерні для гладеньких м’язів внутрішніх органів та поперечносмугастих м’язів серця, що пояснюється тривалим рефрактивним періодом. Тетанічні скорочення, в порівнянні з поодинокими, більш тривалі та сильні. Тетанус викликається сумацією поодиноких м’язових скорочень, коли повторний нервовий імпульс надходить до м’яза раніше, ніж повністю завершиться його поодиноке скорочення (мал. 14). За таких умов відбувається накладання однієї хвилі поодиноких скорочень на другу (явище суперпозиції). Якщо ж кожний наступний імпульс надходить до м’яза в той момент, коли він починає розслаблюватись, то виникає зубчастий неповний тетанус; якщо ж повторний імпульс приходить в період його скорочення – виникає суцільний (повний) або гладенький тетанус. Суцільний тетанус дає більш сильне скорочення, ніж зубчастий тетанус. Частота імпульсів, яка необхідна для виникнення тетанусу, обернено пропорційна швидкості поодиноких скорочень. Для виникнення суцільного тетанусу в повільних РО достатня частота імпульсів 25 за 1 с, а для швидких РО-50 імпульсів за 1 с і більше.
Мал. 14. Формування тетануса в залежності від частоти подразнення
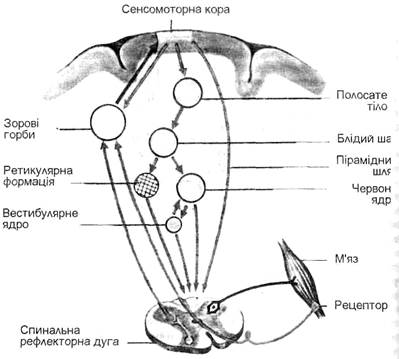
В цілісних м’язах, які складаються з багатьох сотень РО, тетанус є наслідком сумації асинхронних позмінних скорочень різних РО. В режимі такого скорочення досягається максимальне скорочення м’яза. Але максимальне скорочення м’яза виникає лише при певній (оптимальній) частоті подразнення; надмірне збільшення частоти імпульсів викликає зниження сили скорочення м’язів. Для нервово-м’язового препарата жаби оптимальна частота подразнень – 40-50 імпульсів за 1 с Зниження сили скорочення м’язів при високій частоті подразнення (більше 120 імпульсів за 1с) є наслідком стійкої деполяризації мембрани, що виникає при сумації постсинаптичних потенціалів. Контрактура. Поряд з тетанічним скороченням м’язів зустрічається ще одна різновидність тривалого скорочення м’язів – контрактура. Контрактура – це локальне і сильне тривале скорочення м’язових волокон. Воно характеризується стійким напруженням (стягуванням) м’яза з сильно сповільненим розслабленням. Контрактура буває природженою (природжене різке обмеження рухомості внаслідок недорозвинутості м’язів і суглобів) і набутою. Набуті контрактури виникають при порушеннях функцій нервової системи, а також в умовах дії надмірного за силою больового або температурного подразника. Професійні контрактури зумовлені тривалою активністю тих груп м’язів, які інтенсивно використовуються в даній професії. Контрактура при стомленні характеризується збільшенням тривалості періоду скорочення та розслаблення м’язів. В даному випадку вона зумовлена змінами обміну речовин у м’язах. 3. Регуляція напруження м’язів Роль спинного і головного мозку в регуляції напруження м’язів. Функціонування опорно-рухового апарата в цілому і регуляція напруження м’язів зокрема здійснюється завдяки нерозривному взаємозв’язку м’язів з окремими відділами центральної нервової системи (спинного мозку, головного мозку, ко ри великих півкуль), з роботою рухового аналізатора. Спинний мозок (CM) – сегментарний відділ ЦНС, розміщений в хребетному каналі від довгастого мозку до другого поперекового хребця, являє собою тяж з центральним каналом, заповненим прозорою спинномозковою рідиною, яка циркулює між ним і шлуночками головного мозку. Від кожного сегмента CM відходить пара спинномозкових нервів, які іннерюують шию, тулуб, верхні і нижні кінцівки. Навколо центрального каналу CM скупчена сіра речовина (тіла нервових клітин), а зовні неї – біла речовина, яка складається з нервових волокон, що утворюють задні, бічні та передні канатики – висхідні і низхідні провідні шляхи. Сіра речовина утворює парні виступи – передні і задні роги, а в грудному і частково в поперековому відділах CM є ще бокові (бічні) роги, в яких знаходяться нижчі центри симпатичної нервової системи. CM виконує рефлекторну і провідникову функції. Рефлекторна функція полягає в тому, що на рівні CM відбувається складна координація діяльності великої кількості іннервованих м’язів. CM під постійним контролем головного мозку забезпечує оптимальну активність м’язів-синергістів і м’язів-антагоністів, здійснює реципрокну іннервацію, організацію перехресних крокуючих рухів, згинальних, розгинальних, тонічних рефлексів, рефлексів відштовхування при зближенні з опорою, рефлексів на розтягування м’язів. Прикладом розтягувальних рефлексів можуть бути тлінний, ліктьовий, п’ятковий (ахілесів) рефлекси (сухожильні рефлекси). Удари, нанесені гумовим молоточком по сухожиллях, викликають короткочасне розтягування м’язів і відповідні рефлекторні рухові реакції – розгинання ноги в колінному суглобі, згинання руки в ліктьовому суглобі, підошвене згинання стопи. Крім терміну "рефлекс розтягнення", широко використову-ється термін "міотатичний рефлекс" скорочення м’язів-розгиначів при значному розтягненні. Більше як 90% всіх нейронів CM складають проміжні (вставні) нейрони або інтернейрони. Проміжні нейрони забезпечують передачу збудження і процеси гальмування. Для згинального руху в суглобі необхідне не тільки скорочення м’язів-згиначів, а й одночасне розслаблення м’язів-розгиначів – процес гальмування. При збудженні центрів розгиначів, навпаки, гальмуються центри згиначів. Таке координаційне взаємовідношення між моторними центрами спинного мозку називається реципрокною іннервацією. Механізм реципрокної іннервації лежить в основі циклічних рухів – ходьби, бігу, плавання та інших локомоцій. Аксони альфа-мотонейронів перед виходом із спинного мозку часто дають одну або декілька відгалужених волокон (колатералей), які закінчуються на клітинах Реншоу. Аксони клітин Реншоу утворюють гальмівні синапси на мотонейронах даного сегменту спинного мозку, вони зменшують надмірно інтенсивне збудження мотонейронів і беруть участь у регуляції підтримання необхідної пози. Гальмування, яке здійснюється за участю клітин Реншоу, називається зворотним гальмуванням. Перші три сегменти шийного відділу CM, які чають зв’язок з іншими відділами спинного мозку і з нервовими центрами стовбура головного мозку, беруть участь у формуванні статичних (познотонічних і зипрямних) рефлексів, тобто в перерозподілі тонусу м’язів, у підтриманні пози тіла в різних положеннях сидіння, стояння тощо).При відхиленні голови в ту чи іншу сторону подразнюються пропріорецептори м’язів шиї і рефлекторно змінюється тонус м’язів-згиначів, м’язів-розгиначів верхніх і нижніх кінцівок і тонус м’язів тулуба. Наприклад, стійка гімнаста на руках полегшується при відхиленні голови назад, оскільки в даному випадку підвищується тонус м’язів-розгиначів спини і рук. Крім рефлекторної функції, спинний мозок виконує провідникову функцію – тобто проводить нервові імпульси по висхідних шляхах (до головного мозку) і низхідних шляхах (від головного мозку). Так пучки Голя і Бурдаха проводять збудження від про-пріорецепторів м’язів і сухожиль, частково від тактильних рецепторів шкіри і від вісцерорецепторів. По волокнах пучка Голя інформація проводиться від нижньої половини тіла, по волокнах пучка Бурдаха – від верхньої. Волокна цих пучків перериваються в синапсах довгастого мозку. Далі шлях другого нейрона, перехрещуючись, досягає таламуса, де імпульси переключаються на третій нейрон, аксон якого вступає в кору великих півкуль головного мозку, в її задню центральну закрутку. Нервові імпульси від пропріорецепторів м’язів, сухожиль і суглобових зв’язок проходять до головного мозку частково по волокнах спинномозочкових трактів Флексіга і Говерса, розташованих у бокових стовпах спинного мозку, частково по волокнах пучків Голля і Бурдаха. Аксони спинно-мозочкових трактів беруть свій початок від нервових клітин, тіла яких розташовані в задніх рогах спинного мозку, і несуть нервові імпульси до мозочка. Отримуючи пропріоре-цептивну інформацію про стан опорно-рухового апарата, мозочок разом з іншими відділами ЦНС, координує рухи тіла. До низхідних провідних шляхів належать пірамідний (кортікоспінальний) трак і екстрапірамідні шляхи – ретикулоспінальний, руброспінальний, вестибулоспінальний тракти. Оскільки кортікоспінальний тракт проводить нервові імпульси від пірамідальних клітин (гігантські пірамідні нейрони Беца), що знаходяться в руховій зоні кори великих півкуль, і проходить через піраміди довгастого мозку, його називають пірамідним трактом. Пірамідні нейрони кори відповідають за довільні (вольові) м’язові скорочення. Вони посилають нервові імпульси до альфа-мотонейронів передніх рогів спинного мозку. Нервові волокна пірамідного тракту йдуть у бокових і передніх стовпах спинного мозку. Ті нервові волокна пірамідного тракту, що направляються в бокові стовпи, в нижній частині довгастого мозку роблять перехрест, тобто переходять на протилежні сторони. Інша частина волокон передніх стовпів іде, не перехрещуючись у довгастому мозку, до передніх рогів спинного мозку і тільки на рівні сегментів, де закінчується шлях волокон, відбувається перехід цих волокон на протилежні сторони. У зв’язку з перехрещенням пірамідних шляхів порушення моторних центрів кори однієї півкулі головного мозку викликає параліч мускулатури протилежної сторони тіла. В тісній взаємодії з кортікоспінальний трактом функціонує кортікобульбарний тракт, який закінчується в ядрах довгастого мозку. В області головного мозку аксони цих трактів віддають колатералі, які закінчуються в ядрах смугастого тіла, проміжного мозку, варолієвого моста, довгастого мозку, у червоному ядрі; в ретикулярній формації мозкового стовбура (мал. 15). Крім того, до червоного ядра і до ретикулярної формації йдуть безпосередньо нервові тракти від тих же кіркових моторних зон, тобто від зон, звідки починаються кортікоспінальний і кортікобульбарний тракти. Всі вище названі низхідні провідні шляхи, за винятком пірамідального тракту, називаються екстрапірамідними шляхами, або екстрапірамідною системою координації рухової діяльності. До екстрапірамідної системи належать руброспінальний і вестибулоспінальпий тракти. Основне призначення руброспінального тракту, тобто тракту, що несе нервові імпульси від червоного ядра до рухових центрів спинного мозку, полягає в управлінні тонусом м’язів і в координації мимовільних рухів. Руброспінальний тракт несе також імпульси від мозочка, ядра вестибулярного нерва, смугастого тіла. В регуляції наруження м’язів важлива роль належить головному мозку, який складається з довгастого мозку, заднього мозку (вароліїв міст і мозочок), середнього мозку, проміжного мозку і великих півкуль. Із основи головного мозку виходять 12 черепномозкових нервів.
Мал. 15. Низхідні впливи на мотонейрон.
Довгастий мозок і вароліїв міст – єдиний структурно-функціональний відділ головного мозку. Він є безпосереднім продовженням спинного мозку. Рефлекси заднього мозку більш складно координовані, ніж рефлекси спинного мозку. Причому в координації ряду рефлекторних актів беруть спільну участь нервові структури довгастого мозку і варолієвого моста. В товщі білої речовини заднього мозку зосереджені численні скупчення сірої речовини, які називаються ядрами і центрами. В області варолієвого моста знаходяться ядра V, VI, VII і VIII пар черепномозкових нервів – ядра трійчастого, відвідного, лицевого і слухового нервів. Ядро вестибулярного нерва – це комплекс ядер органа рівноваги: ядро Швальбе, ядро Бехтерева, ядро Дейтерса. Ядро Дейтерса разом з ядрами, що сприймають нервові імпульси від пропріорецепторів шиї, бере безпосередню участь у формуванні статичних рефлексів – познотонічних та установчих. У задньому мозку існують групи нейронів, які підвищують тонус м’язів-розгиначів. Основними структурно-функціональними компонентами середнього мозку є чотиригорбикове тіло, червоне ядро, чорна субстанція, ядра III і IV пар черепномозкових нервів, ніжки мозку, частина ретикулярної формації. В передніх горбиках чотиригорбикового тіла розміщені первинні підкіркові зорові центри, в задніх – первинні підкіркові слухові центри. Вони беруть участь у формуванні зорових і слухових орієнтувальних рефлекторних реакцій у відповідь на несподівані, раптові світлові і звукові подразнення, що має важливе значення в утворенні умовних рефлексів і в мобілізації уваги дітей. Червоні ядра (їх два) підвищують тонус м’язів-розгиначів. В них переключаються нервові імпульси від волокон екстрапірамідної системи і мозочка. Екстрапірамідні волокна направляються сюди від базальних гангліїв кінцевого мозку. Далі імпульси по руброспінальному тракту йдуть до передніх рогів спинного мозку, збуджуючи альфа- і гамма-мотонейрони м’язів-згиначів. Чорна субстанція (скупчення нервових темнозабарвлених клітин) має відношення до регуляції складних актів жування і ковтання, а також тонких рухів пальців. Важлива роль в регуляції напруження м’язів належить мозочку, який має аферентні і еферентні зв’язки з усіма руховими центрами. До мозочка по аферентних шляхах надходить інформація від: 1) пропріорецепторів по спинно-мозочкових висхідних шляхах; 2) вестибулярних ядер заднього мозку; 3) усіх рухових зон кори великих півкуль – через варолієвий міст. По еферентних шляхах від мозочка посилаються нервові імпульси до кори великих півкуль головного мозку (через таламус), вестибулярних ядер заднього мозку, червоного ядра, ретикулярної формації довгастого мозку і варолієвого моста. Основне функціональне значення мозочка полягає в доповненні і корекції діяльності рухових центрів кори і стовбурної частини головного мозку. Він узгоджує швидкі і повільні рухи, регулює позу м’язовий тонус. Мозочок бере участь у координації всіх складних рухових актів організму, включаючи довільні рухи. В літературі (Р.Шмідт, 1985) висловлюється думка про те, що інформація про замисел руху передається корою великих півкуль головно: мозку в мозочок для перетворення в програму руху, яка посилається зворотно через таламус до рухових областей кори. Після цього стає можливим здійснення руху. У тварин з вирізаним мозочком спостерігали зниження м’язового тонусу (атонія), швидку втому (астенія), мимовільні рухи кінцівок і голови (астазія) і порушення координації рухів (атаксія). Спостереження, проведені над людьми, з паталогічним ураженням мозочка, дозволили вченим відмітити такі характерні симптоми, як ністагм, головокружіння, тремор, дефекти мови. Мозочок не тільки регулює діяльність скелетних м’язів, а й впливає на діяльність внутрішніх органів, на рівень артеріального тиску і склад крові. Довільна рухова діяльність людини не можлива без участі кори великих півкуль головного мозку. Кора – це сіра речовина на поверхні великих півкуль, скупчення величезної кількості нервових клітин. При загальному огляді кори в плані її розвитку і будови розрізняють нову кору і стару кору. Нова кора знаходиться на верхньопередній, задній і боковій поверхні півкуль. Стара кора розміщена на нижній і внутрішній поверхнях півкуль. Три основні найбільші борозни – центральна, бокова, тім’яно-потилична ділять нову кору на чотири долі або частки: лобову, тім’яну, потиличну і скроневу. Менші, ніж основні, борозни розмежовують закрутки. Так, у лобовій частці в області нової кори розрізняють верхню, середню і нижню закрутки, а поруч з ними перпендикулярно їм розташована передня центральна закрутка, яка відділяється центральною борозною від задньої центральної закрутки, що відноситься до тім’яної долі. На боковій поверхні скроневої частки теж розрізняють верхню, середню і нижню закрутки. Передня центральна закрутка являє собою первинну моторну зону. Локалізація в цій зоні рухових точок, від яких посилаються сформовані у вертикальних колонках нервові імпульси до скелетних м’язів, відповідає послідовності представництва рецепторних полів у задній центральній закрутці (перехрестя: права півкуля – ліва сторона тіла; ліва півкуля – права сторона тіла; верх-низ; низ-верх). Пірамідні клітини моторної зони (гігантські піраміди Беца), що входять до складу вертикальної колонки як структурно-лункціональної одиниці кори, посилають імпульси до мотонейронів, які іннервують скелетні м’язи і відповідають за довільні м’язові скорочення. Кіркова регуляція рухової діяльності не обмежується функцією первинної моторної зони. Поруч з нею в лобовій частці і за її межами існують вторинні і третинні моторні зони, які формують складні рухові акти за участю базальних гангліїв, мозочка і структур екстрапірамідної системи. В третинних зонах лобової частки здійснюється свідоме програмування довільних рухів,, визначення мети поведінки, рухових задач. Проекція рецепторних полів нижньої частини тіла знаходиться у верхній частині задньої центральної закрутки кори мозку, а верхніх частин тіла -з нижній частині закрутки, тобто схема соматосенсорної проекції виглядає так: низ-верх, верх-низ, ліва половина тіла – права півкуля головного мозку, права половина тіла – ліва півкуля. Представництво (проекція) сенсорної функції в задній нейтральній закрутці відповідає представництву рухової функції різних частин тіла в передній центральній закрутці. При подразненні поверхні шкіри короткими точковими доторками первинна відповідь у соматосенсорній зоні кори локалізується в суворо обмеженій ділянці. Методом порівняння точок подразнення на поверхні шкіри з точками кори, які дають відповідь, вченими виявлені проекції різних областей тіла на соматосенсорну зону кори (SI) "точка в точку". Таку локалізацію називають соматотопією, або топографічним представництвом. Найбільш обширні ділянки в корі виявлені для губ, обличчя і кистей рук. У соматосенсорні зони задньої центральної закрутки, а звідси в передню центральну закрутку (рухова зона) надходять потоки нервових імпульсів, викликані збудженням пропріорецепторів м’язів, сухожиль, суглобів. Обширне представництво кистей рук у корі великих півкуль головного мозку, тісний зв’язок цієї області кори з мовними та іншими центрами, могутній вплив чутливості і рухової діяльності пальців рук на функціональний стан всього організму повинні враховувати педагоги при навчанні і вихованні дітей. Роль рухового аналізатора в регуляції напруження м’язів. Руховий аналізатор забезпечує організм інформацією про положення тіла в просторі, про міру скорочення кожного м’яза, бере безпосередню участь у координації рухів і регулює тонус м’язів. Периферійний відділ цього аналізатора являє собою пропріорецептори м’язів, сухожиль, суглобів (мал. 16, 17).
Мал. 16. М’язові рецептори.
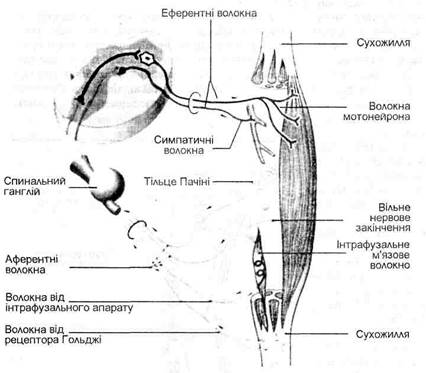
Термін "пропріорецептор" походить від латинського слова "пропріус", що означає власний. В скелетних м’язах знаходяться пропріорецептори, які називаються м’язовими веретенами, а в сухожиллях на місці з’єднання з м’язовою тканиною – сухожильні органи Гольджі (мал. 18). Суглоби мають декілька видів пропрюрецепторів – рецептори, що нагадують структуру тілець Пачіні, сухожильних органів Гольджі і тілець Руффіні. Тут виявляються також вільні нервові закінчення. Пропріо-рецептори суглобів сигналізують про положення частин тіла (кінцівок), напрям і швидкість їхнього руху, м’язову силу. Адекватним подразненням для м’язових веретен і сухожильних органів Гольджі є їх розтягнення.
Мал. 17. Іннервація м’язів.
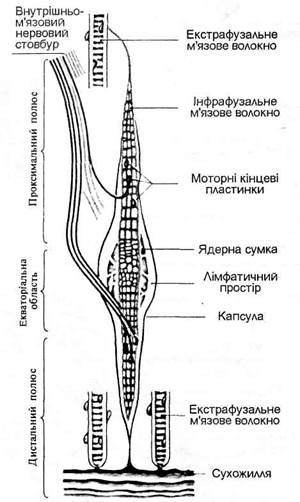
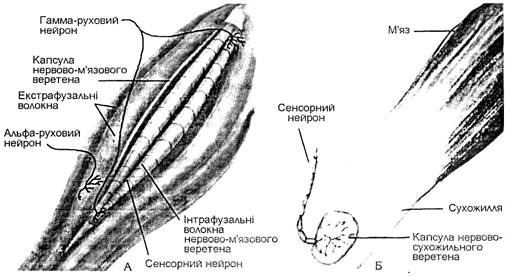
М’язові веретена розташовані в скелетних м’язах між поперечносмугастими м’язовими волокнами, прикріпляючись до них паралельно. Кожне м’язове веретено має сполучнотканинну капсулу, всередині якої розміщені інтрафузальні м’язові волокна. Звичайні м’язові волокна основної маси м’язів називаються екстрафузальними, вони виконують рухову функцію. Інтрафузальні м’язові волокна менші за розміром від екстрафузальних волокон. Їх основною функцією є рефлекторна регуляція (через спинний мозок) діяльності екстрафузальних м’язових волокон. У людини існують два типи інтрафузальних м’язових волокон, які розрізняють за розташуванням ядер. У м’язових волокнах обох типів усі ядра зосереджені в середній частині волокон. Ядра м’язових волокон першого типу розташовані у вигляді тісного скупчення (ядерна сумка), яке займає на короткій відстані в середній частині всю товщу волокна. Ядра м’язових волокон другого типу розташовані в один ряд у вигляді ланцюжка. Інтрафузальні м’язові волокна з ядерною сумкою вдвічі товщі і майже вдвічі довші, ніж інтрафузальні волокна з ядерним ланцюжком.
Мал. 18. Нервово-м’язове веретено (А) і нервово-сухожильне веретено (Б).
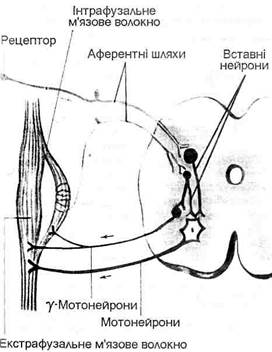
Інтрафузальні м’язові волокна (їх по декілька в кожному м’язовому веретені) мають периферичні полярні скоротливі ділянки. Середні частини м’язових волокон і м’язового веретена в цілому не скоротливі, а тільки можуть розтягуватись при розслабленні і розтягуванні екстрафузальних м’язових золокон і м’яза в цілому або при скороченні полярних частин інтрафузальних м’язових волокон. До скоротливих ділянок інтрафузальних м’язових волокон обох типів підходять від передніх рогів спинного мозку рухові нервові волокна гамма-мотонейронів. Екстрафузальні м’язові волокна іннервуються альфа-мотонейронами, тіла яких також знаходяться в передніх рогах спинного мозку. Гамма-волокна утворюють два типи закінчень на периферичних (полярних) ділянках інтрафузальних м’язових волокон: гамма-пластинки на інтрафузальних волокнах з ядерною сумкою і гамма-кущовидні закінчення на інтрафузальних волокнах з ядерним ланцюжком. В кожне м’язове веретено в ділянці його ядерної зони проникає найбільшої в організмі товщини чутливе мієлінове нервове волокно групи А-альфа, яке всередині веретена ділиться на гілки. Ці гілки у вигляді спіралеподібних нервових закінчень обвивають середині частини всіх інтрафузальних м’язових волокон даного веретена. Це первинні чутливі закінчення обох типів інтрафуззальних м’язових волокон. На відміну від інтрафузальних м’язових волокон з ядерною сумкою інтрафузальні м’язові волокна з ядерним ланцюжком іннервуються додатково вторинними чутливими закінченнями аферентного волокна меншої товщини порівняно з товщиною волокон, що дають первинні чутливі закінчення.
Мал. 19. Саморегуляція тонусу м ‘язів. Первинні закінчення інформують про зміну розтягування м’язів і швидкість його подовження в динамічній фазі. Вторинні закінчення реагують на постійне натягнення м’яза і спеціалізовані для передачі інформації про положення м’яза. Вони мало чутливі до швидкості розтягнення. Ця їх властивість пов’язана з структурно-функціональними особливостями інтрафузальних м’язових волокон з ядерним ланцюжком, які іннервуються не динамічними, а статичними гамма-волокнами гамма-мотонейронів (мал. 19). Роздільна гамма-іннервація інтрафузальних волокон з ядерною сумкою і з ядерним ланцюжком робить можливою роздільну регуляцію динамічної і статичної діяльності м’язів. Ітрафузальні м’язові волокна з ядерною сумкою, які мають первинні аферентні нервові закінчення, активно реагують на динамічне розтягнення м’яза. При цьому різко збуджуються первинні нервові закінчення, нервові імпульси від яких надходять у спинний мозок і моносинаптично переключаються на альфа-мотонейрони передніх рогів спинного мозку. Альфа-мотонейрони посилають нервові імпульси до екстрафузальних волокон даного м’яза. Отже, вслід за розслабленням і розтягненням м’язів настає їх рефлекторне скорочення, підтримується необхідний тонус (механізм регуляції довжини м’язів). Таке рефлекторне підтримування довжини м’язів має особливо важливе значення для сталості тонусу мускулатури, яка забезпечує збереження пози. Якщо в людини у вертикальному положенні відбувається таке незначне згинання в колінному суглобі, яке вона не може ні відчути, ні побачити, то в результаті розтягнення чотириголового м’яза стегна збільшується імпульсація від первинних чутливих закінчень веретен, закладених у цьому м’язі, збільшується збудження альфа-мотонейронів чотириголового м’яза, підвищується його тонус і миттєво зупиняється згинання в суглобі, яке перед цим почалось. Так попереджується порушення пози стояння. Навпаки, при надмірному скороченні м’яза ступінь збудження закладених у ньому м’язових веретен зменшується. Таким чином, дана регуляторна система сприяє збереженню постійної довжини м’яза. Необхідний рівень чутливості м’язових веретен підтримується скоротливою діяльністю інтрафузальних м’язових волокон. При моносинаптичному переключенні нервових імпульсів з чутливих волокон, що йдуть від м’язових веретен даного м’яза на альфа-мотонейрони, які збуджуються, одночасно гальмуються альфа-мотонейрони м’язів-антагоністів. Первинні чутливі нервові закінчення м’язових веретен можуть збуджуватись під впливом розтягнення м’яза, а також в результаті скорочення інтрафузальних м’язових волокон при збудженні фузімоторних гамма-мотонейронів. Отже, існують два механізми збудження м’язових веретен: 1) розтягнення м’яза і 2) скорочення м’язових волокон. Завдяки наявності механізму скорочення інтрафузальних м’язових волокон, м’язові веретена продовжують виконувати функцію датчиків довжини м’язів навіть при скороченні екстрафузальних волокон. Сухожильні органи Гольджі (тільця Гольджі) – пропріорецептори, які знаходяться в ділянках сухожиль, що примикають до м’язів. Вони з’єднуються з м’язовими екстрафузальними волокнами послідовно і тому розтягуються і збуджуються не при розтягненні м’яза, як м’язові веретена, а, навпаки, при скороченні м’яза. Сухожильний пропріорецептор (сухожильний орган Гольджі) складається з сухожильних ниток, які відходять приблизно від десяти екстрафузальних м’язових волокон, і оточений сполучнотканинною капсулою. Довжина пропріорецептора може досягати 1 мм. Від сухожильних органів Гольджі відходять і направляються в спинний мозок чутливі нервові волокна, які мають набагато менший діаметр, ніж діаметр волокон, що відходять від м’язових веретен. В спинному мозку нервові волокна, що відходять від органів Гольджі, активують гальмівні вставні нейрони, які пригнічують активність альфа-мотонейронів власного м’яза і викликають збудження альфа-мотонейронів м’яза-антагоніста. В загальному м’язові веретена змушують власний м’яз скоротитися після його розслаблення, а сухожильні органи Гольджі – розслабитись після скорочення. Рефлекси, які виникають при збудженні сухожильних органів Гольджі, сприяють підтриманню сталості напруження м’язів. В регуляції діяльності кожного м’яза беруть участь дві регуляторні системи: система регуляції довжини, роль датчика в якій виконують м’язові веретена, і система регуляції напруження, датчиками якої служать сухожильні органи Гольджі. Провідниковий і центральний відділи рухового аналізатора. Аферентні нервові волокна від м’язових веретен, сухожильних органів Гольджі і суглобових рецепторів після входження через задні корінці в спинний мозок зразу ж переходять у склад білої речовини задніх канатиків або стовпів, по яких піднімаються до першого синаптичного переключення в ядрах довгастого мозку (ядра пучків Голля і Бурдаха). Пучки Голля і Бурдаха розташовані по боках задньої серединної борозни. Від ядер цих пучків у довгастому мозку направляються волокна других нейронів до ядер таламуса, від ядер таламуса волокна третіх нейронів піднімаються до задньої центральної закрутки, де знаходиться кіркова соматосенсорна зона.
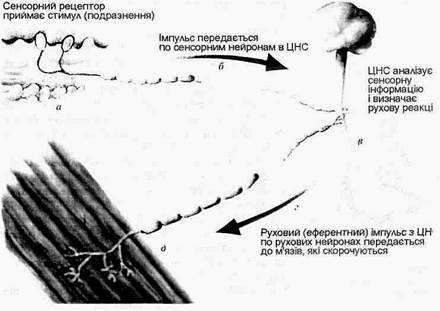
Мал.20. Послідовність процесів в сенсорно-руховій інтеграції (Дж.Уілмор, Д.Костілл, 1997).
Таким чином, провідниковий відділ рухового аналізатора, його висхідний шлях співпадає з висхідним шляхом тактильної чутливості аналізатора шкірного чуття. Проте руховий аналізатор має також представництво в передній центральній закрутці (рухова зона) і посилає інформацію від своїх пропріорецепторів до мозочка, який бере участь у координації рухів. Загальна схема послідовності процесів сенсорно-рухової інтеграції подана на мал.20. Імпульси, які надходять в ЦНС від пропріорецепторів рухового аналізатора, суттєво впливають на функції різних органів і систем організму. М’язова робота викликає підсилення функцій серцево-судинної, дихальної, видільної та інших систем. Після її закінчення підвищується потреба в їжі і посилюється робота травних органів. При м’язовій роботі встановлюються зв’язки між відповідними центрами рухового аналізатора і центрами, що регулюють роботу внутрішніх органів. Систематична рухова діяльність стимулює розвиток рухового аналізатора, сприяє його функціональному удосконаленню. Тому спортсмени краще орієнтуються в просторі, точніше координують свої рухи в часі і просторі, точніше, ніж неспортсмени, диференціюють м’язові зусилля. Пропріорецептори рухового аналізатора, як і рецептори шкіри, ускладнюються і функціонально удосконалюються протягом тривалого періоду постнатального життя, досягаючи остаточної зрілості в 12-15 років. На 3-му – 4-му тижні після народження дитини виникають умовні рефлекси на пропріорецептивні подразнення. В подальшому виникають все нові і нові умовні рефлекси, пов’язані з діяльністю рухового аналізатора. Формується велика кількість умовних рефлексів II роду – такі званих оперантних або інструментальних, умовних рефлексів, які лежать в основі вироблення різноманітних рухових, професійних навичок. При м’язовій діяльності виникають зв’язки між центрами рухового аналізатора і центрами, що регулюють діяльність різних органів, у тому числі органів системи кровообігу, дихання, травлення, виділення. Встановлюється тонке узгодження між роботою рухового апарата і внутрішніх органів. Відомо, що будь-яка м’язова робота стимулює насамперед систему енергозабезпечення. А така стимуляція без надходження прорпріорецепторної інформації неможлива. Суттєво впливає руховий аналізатор на розвиток моторної мови дітей (М.М.Кольцова, 1975, та ін). Цьому сприяє близькість рухової проекційної зони кори головного мозку до центрів умовних рефлексів і обширна рухова проекція кисті руки. Виявлена висока кореляція рівня розвитку мови і точних високо-координованих рухів пальців рук. 4. Фізіологічні механізми внутрішньом’язової і міжм’язової координації функцій опорно-рухового апарата. Електроміограма Регуляція напруження м’язів здійснюється через механізми управління м’язовим апаратом – механізми внутрішньом’язової і міжм’язової координації. Внутрішньом’язова координація рівня напруження даного м’яза досягається: 1) регуляцією числа активних РО даного м’яза; 2) режимом активності РО (роботи більшості РО в тетанічному режимі); 3) одночасною активністю більшості мотонейронів. РО, яка в даний момент часу одержує від мотонейрона нервові імпульси і її волокна скорочуються, називається активною РО. Чим більше активних РО у даного м’яза, тим більше напруження він розвиває. Число активних РО визначається інтенсивністю збуджуючих еферентних впливів ЦНС, які в значній мірі регулюються вольовими зусиллями. Якщо для виконання даного рухового завдання м’яз повинен розвивати невелике напруження, то до його мотонейронів (звичайно малих мотонейронів з низьким порогом збудження) надходять відносно слабкі за силою збуджуючі нервові імпульси, які активують повільні РО. Для розвитку м’язом великого напруження ЦНС активуються великі високопорогові мотонейрони, які зключають в діяльність швидкі (великі) РО. Цей механізм включення (рекрутування) РО у відповідності з їх розмірами називається "правилом розміру" Е.Хеннеман). У відповідності з цим правилом найменші (повільні) РО активні при будь-якому напруженні м’яза, а великі (швидкі) РО активні лише при сильному м’язовому напруженні. Тому степінь використання, а отже і функціонального вдосконалення тренування) великих швидких РО у звичайних умовах повсякденної діяльності завжди нижчий, ніж малих повільних РО. Активність різних видів РО визначається потужністю і тривалістю виконуваної роботи. При виконанні тривалої м’язової роботи помірної потужності в першу чергу функціонують низькопорогові повільні РО. В міру їх стомлення в роботу включаються більш зисокопорогові РО. Режим активності РО. Режим роботи РО визначається частотою імпульсації мотанейронів. Чим вища (в межах оптимуму частоти) частота імпульсації мотонейрона, тим більше напруження розвиває РО перехід зубчастого тетанусу в гладенький). Частота імпульсації мотонейронів визначається збуджуючими впливами рухових центрів кори мозку і підкіркових моторних центрів. Функціональний стан нервових центрів, в свою чергу, залежить від вольових зусиль даної людини та емоцій. Якщо частота імпульсації з боку ЦНС невелика, то працюють низькопорогові, малі (повільні) мотонейрони, які активують повільні РО. А оскільки частота імпульсації мотонейронів невелика, то РО працюють у режимі поодиноких скорочень або в режимі зубчатого тетанусу. Такий режим роботи РО забезпечує виконання м’язом тривалої, малоінтенсивної невтомливої роботи, направленої, наприклад, на збереження пози тіла. При більш високій частоті імпульсації з боку ЦНС, раніше працюючі в режимі зубчатого тетанусу, низькопорогові РО починають працювати в режимі гладенького тетанусу. У цей період, спочатку в режимі поодиноких скорочень, а потім у тетанічному режимі, починають працювати і великі, високопорогові РО. Такий режим роботи м’язів пов’язаний з великими витратами енергії, він дуже втомливий, а тому короткотривалий. Зв’язок активності РО у часі. Ступінь напруження м’яза визначається також тим, як пов’язані в часі імпульси, які надходять від різних мотонейронів до даного м’яза. В звичайних умовах більшість РО даного м’яза працює асинхронно-рівномірно, незалежно одна від одної, що зменшує амплітуду фізіологічного тремору і забезпечує необхідну плавність скорочення м’яза та велику точність утримання необхідної пози. Втома м’яза приводить до синхронного (одночасного) скорочення його РО. При цьому рух втрачає плавність, порушується точність, виникає втома – великі коливальні рухи з частотою біля шести коливань за 1 с. При роботі РО у режимі гладенького тетанусу характер зв’язку в час імпульсної активності мотонейронів практично не впливає на величину максимального напруження. Але при короткочасних скороченнях або на початку будь-якого скорочення м’язів синхронізація імпульсної активності мотонейронів впливає на швидкість розвитку напруження ("градієнт сили" або "вибухова сила"): чим більше співпадань у скоротливих циклах різних РО на початку скорочення м’яза, тим швидше зростає величина напруження. Основою механізмів міжм’язової координації є координація активності ряду м’язів шляхом включення «потрібних» для успішного виконання даної вправи м’язів-синергістів і виключення «непотрібних» в даний момент м’язів-антогоністів. Завдяки високій досконалості механізмів міжм’язової координації істотно зменшується загальна напруженість м’язів, зростає їх максимальна довільна сила. Електроміограма. У відповідності до частоти подразнень збуджене м’язове волокно відповідає повним ритмом електричних потенціалів. З допомогою електроміографа ці електропотенціали м’язових волокон реєструють у вигляді електроміограми (ЕМГ). При слабкому скороченні м’яза, коли активні тільки деякі його РО, вдається зареєструвати електричну активність окремих РО. При збудженні багатьох м’язових волокон відбувається сумація їхніх струмів дії, яка має характер осциляції – потенціали дії активних РО нашаровуються один на одного (явище інтерференції). При цьому амплітуда коливань збільшується, а ЕМГ перетворюється у інтерференційну ЕМГ, в якій не вдається відокремити ПД окремих РО. Між сумарною електричною активністю і силою скорочення м’яза в межах від слабких до субмаксимальних фізичних навантажень існує пряма залежність. Для кількісної оцінки електричної активності м’язів проводять інтегрування інтерференційної ЕМГ- визначення загальної площі кривої коливань електричної активності досліджуваного м’яза. В міру розвитку втоми амплітуда ЕМГ збільшується, що пояснюється включенням в роботу високопорогових РО. За ЕМГ оцінюють рівень вдосконалення рухової навички (правильність включення окремих м’язів), структуру рухової навички (черговість включення окремих м’язів),напруженість м’язів, вираженість втоми тощо. 5. Робота м’язів і її механічна ефективність Скорочуючись м’яз виконує зовнішню механічну роботу – добуток величини сили (наприклад, піднятого вантажу) на відстань переміщення. Одиниця виміру роботи – кілограмометр (кгм) або джоуль (Дж). Один Дж – це робота сили в 1 ньютон при переміщенні тіла на відстань 1 м. Відношення величини роботи до часу її виконання називається потужністю. Потужність вимірюється в ватах (Вт), кгм/с або кгм/хв. (1 Дж/с = 1 Вт; 1 кгм/с = 9,8 Вт; 1 Вт = 0,102 кгм/с або 6,12 кгм/хв.) Ефективність роботи м’язів (ефективність використання енергії) оцінюють шляхом визначення коефіцієнта корисної дії (ККД). ККД – це виражене в процентах відношення корисної механічної енергії (КМЕ), затраченої на роботу, до загальних енерговит-рат (ЗЕ) мінус витрати енергії у стані спокою за період виконання роботи (ВЕС).
Величина ККД при ходьбі – 20-25%, бігу, їзді на велосипеді, греблі, штовханні ядра – 30-40%, підніманні штанги – 10-15%, плаванні – 3-6%. Низький ККД при плаванні пояснюється значними енерговитратами, пов’язаними з високою теплоємністю води. Таким чином, тільки 30-40% енергії скорочення м’язів витрачається на механічну роботу, а решта 60-70% розсіюється у вигляді тепла (для порівняння: ККД паровоза – біля 6%, тепловоза – 40%, сучасного електровоза – 60%, стрибаючого кенгуру – 76%). Висока економічність руху кенгуру зумовлена здатністю її скелетних м’язів багаторазово використовувати вивільнену енергію, яка нагромаджується в "м’язах-пружинах". Явище повторного використання енергії (рекуперація) спостерігається і у спортсменів – стрибунів, лижників, гребців. Оскільки потенційна енергія м’язів може зберігатися дуже короткий час – (через 5-6 с вона майже повністю розсіюється), її використання можливе тільки при великому темпі рухів. Принцип рекуперації враховується при конструюванні спеціальних черевиків з пружинами для стрибання "покенгуриному". Закон середніх навантажень і середніх швидкостей. Ефективність роботи м’язів залежить від величини навантаження і швидкості скорочення – він найбільший при навантаженнях, які складають половину від максимальної сили м’язів (закон середніх навантажень), і при швидкості скорочення – 35% від максимальної (закон середніх швидкостей). При більш високих швидкостях значна частина енерії витрачається на подолання внутрішнього тертя (в’язкості): чим більша швидкість скорочення м’язів тим більше внутрішнє тертя. Закони середніх навантажень і середніх швидкостей необхідно враховувати на виробництві і в практиці фізичного виховання. У процесі тренувань юних спортсменів одночасно із збільшенням функціональних можливостей рухового апарата збільшуються абсолютні значення середніх навантажень і швидкостей. Тому величину середніх навантажень і швидкостей в кожному окремому випадку необхідно визначати експериментально, враховуючи вік, розвиток окремих груп м’язів та інші фактори. Прикладом практичного використання законів середніх швидкостей і середніх навантажень може бути їзда на велосипеді з переключенням передачі або зигзагоподібне піднімання туристів вгору тощо. При вантажах більших від величини напруження, яку м’яз розвиває, зовнішня робота дорівнює нулю. Однак у цьому випадку м’яз знаходиться в ста напруження і витрачає енергію (статична робота), яка повністю перетворюється в тепло. У спортивній практиці механічну продуктивність роботи (ККД м’язової діяльності) розраховують за формулою:
де: MP – механічна (зовнішня) робота в кгм Сn02 – обсяг використаного організмом при роботі кисню в мл; 0,49 – коефіцієнт еквівалентності між механічною роботою і обсягом використаного кисню Із зростанням натренованості спортсмена ефективність роботи м’язів зростає. Це досягається шляхом зменшення енерговитрат за рахунок покращення: діяльності (економічності функціонування) дихальної і серцево-судинної систем, які забезпечують працюючі м’язи киснем та енергосубстратами, а також за вдяки покращенню координації рухів (гальмування активності м’язових груп, які не беруть участі у ви конанні даної роботи). Динамічна і статична робота. М’язова робота при якій відбувається переміщення вантажу і рух кісток в суглобах, називається динамічною. При виконанні такої роботи спостерігається вкорочення і розслаблення м’язів. Робота, при якій м’язові волокна розвивають напруження, але майже не вкорочуються називається статичною (м’язи знаходяться в ізометричному напруженні). Динамічна робота може продовжуватись тривалий час. Це пояснюється почерговим скороченням і розслабленням окремих м’язів. Динамічна робота м’язів лежить в основі ходьби, бігу, плавання, греблі спортивних ігор, багаточисельних гімнастичник вправ тощо. Статична робота швидко викликає стомлення і не може продовжуватись тривалий час. Це пояснюється розвитком у рухових центрах працюючих м’язів захисного гальмування, яке завжди виникає тим швидке, чим інтенсивніша пропріорецептивна імпульсація. В умовах інтенсивних статичних навантажень, внаслідок розвитку великого внутрішньом’язового тиску, порушується капілярний кровообіг м’язів, розвивається гіпоксемія, нагромаджується значна кількість продуктів обміну. Все це призводить до швидкого розвитку втоми і мимовільного припинення статичного зусилля. Натуженим. Статичні вправи силового характеру виконуються з затримкою дихання і натуженням. Натуження пов’язане з напруженням м’язів живота та діафрагми при підніманні вантажу (у штанговому спорті) або при подоланні опору зовнішньої опори (в єдиноборствах). При цьому різко підвищується внутрішньочеревний та грудний тиск, фіксується грудна клітка, знижується забезпечення організму киснем. Натуження завжди починається на вдиху, закінчується на видиху. Досить часто натуження поєднується з напруженням м’язів кінцівок. Різке зменшення відтоку венозної крові до правого передсердя при натуженні призводить до застою крові у венозній частині великого кола кровообігу, при цьому зменшується і надходження крові до лівого передсердя. Як наслідок, зменшується величина систолічного обсягу крові та падає кров’яний тиск (у натренованих людей звуження периферичних судин може призвести до незначного підвищення кров’яного тиску). Після закінчення натуження, внаслідок збільшення притоку крові до серця, збільшується систолічний обсяг крові і підвищується кров’яний тиск. При натуженні відзначають незначне зростання м’язової сили, що, ймовірно, є наслідком подразнення механо- та хеморецепторів, розташованих у легеневій тачеревній порожнинах. Ці рецепторні впливи через ЦН С підвищують збудливість м’язів. Не виключено тут формування домінанти, яка забезпечує посилення збудливості працюючих рухових центрів кори мозку за рахунок імпульсів, адресованих іншим нервовим центрам. При виконанні статичних і швидкісно-силових вправ найбільше зусилля розвивається при затриманні дихання, найменше – при вдиху. Особливо помітно натуження проявляється у ненатренованих осіб. З ростом спортивної майстерності вираженість його прояву знижується. Це пояснюється включенням дихального акту у систему умовнорефлекторних зв’язків конкретної рухової навички. Вплив статичних навантажень на організм людини вивчено недостатньо, що значно ускладнює Використання статичних вправ в оздоровчому тренуванні школярів і дорослих. Статичні навантаження повинні бути адекватними позам і статичним напруженням, які властиві дітям і підліткам в повсякденному житті і при заняттях фізкультурою та спортом (І. В.Павлова, 1986). Найбільш адекватними статичними навантаженнями є зусилля, направлені на утримання маси тіла в положенні лежачи на стегнах, лежачи на спині, верхній чи нижній половині свого тіла. При цьому спостерігається тривале (до відмови) напруженням великих груп м’язів. Розвиваючи статичну витривалість, такі вправи сприяють удосконаленню вегетативного забезпечення м’язових напружень за рахунок оптимізації функцій серцево-судинної і дихальної систем, вдосконалення механізмів аеробного енергозабезпечення м’язової діяльності. Статичні навантаження (з вихідним положенням стоячи) не повинні виконуватись до відмови і перевищувати 70% від максимального зусилля. Тривалість статичних напружень великих груп м’язів в положенні лежачи на стегнах з утриманням ваги тіла до відмови у 10-річних підлітків складає 89,33 + 6,11 с, у 15-річних – 100,3 + 8,95 с (Л.Г.Петрова, В.Г.Петров, 1984). Нами встановлено ряд особливостей статичної витривалості у гімнастів і легкоатлетів в умовах виконання статичних напружень (утримання ніг, лежачи на спині під кутом 45°. і утримання тулуба на гімнастичній лаві, "висячи" на стегнах). Більш висока статична витривалість гімнастів (107 і 79,5 с.) в порівнянні з легкоатлетами (96,0 і 62,8 с.) пояснюється тим, що в тренувальних програмах гімнастів більше статичних вправ (утримання фіксованої пози), ніж у легкоатлетів. Тривалість же відновного періоду була значно меншою у легкоатлетів (176 ± 2,8 с), ніж у гімнастів (204 ± 3,0 с, Р<0,001). Важливо відмітити, що тривалість утримання ніг під кутом 45° (як у гімнастів, так і у легкоатлетів більша, ніж тривалість утримання тулуба на гімнастичній лаві (відповідно на 34% і 25%). Різницю одержаних показників можна пояснити більш природним положенням тіла при утриманні ніг і меншою вагою нижніх кінцівок в порівнянні з вагою верхньої половини тіла. Статичні зусилля з напруженням великих груп м’язів при утриманні власного тіла в фіксованих статичних позах є найбільш адекватними для натренованих осіб, вони пропонуються також для широкого застосування в практиці фізичного тренування школярів і дорослих. При тренуванні статичної витривалості доцільно поєднувати ізометричні скорочення м’язів (не тривалих і не максимальних за потужністю) з динамічними, з включенням вправ на розслаблення. Оптимальним навантаженням для розвитку статичної витривалості є статичні вправи тривалістю 80-85% часу від максимально можливого утримання статичного напруження. 2.4. ПРАКТИЧНІ ЗАВДАННЯ Матеріальне забезпечення заняття. Таблиці, схеми, діапозитиви (кодопозитиви) з питань даної теми, діапроектор, кодоскоп. Осцилограф, підсилювач змінного струму, нашкірні електроди, лейкопластир, кистьовий динамометр з рухливою стрілкою, міотонометр Козловського або інші. РОБОТА 1 Дата добавления: 2015-11-26 | Просмотры: 3524 | Нарушение авторских прав |