|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
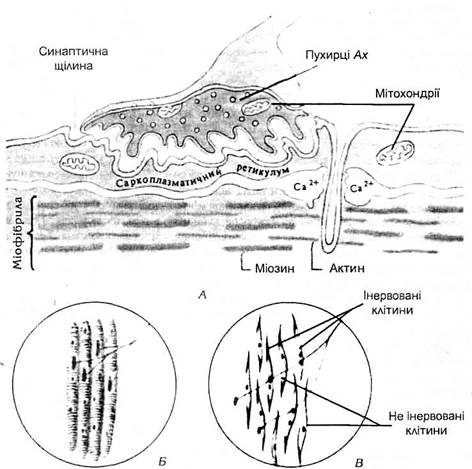
Нервово-м’язове з’єднання. Механізм м’язового скороченняЗбудження і скорочення скелетного м’яза викликається нервовими імпульсами, які надходять від мото-нейронів спинного мозку або рухових ядер головного мозку до нервово-м’язових синапсів. Нервово-м’язові синапси відрізняються синаптичними складками, завдяки яким значно збільшується площа синаптичноі щілини. Синапс складається з мембрани нервового закінчення (пресинаптична мембрана), мембрани м’язового волокна (постсинаптична мембрана) і розміщеної між ними синаптичної щілини (мал.6). В пресинаптичному нервовому закінченні є багато міхурців, які містять медіатор ацетилхолін (АХ). Вихід ацетилхоліну з пресинаптичних міхурців у синаптичну щілину відбувається під впливом нервових імпульсів. Переходячи через синаптичну щілину, ацетилхолін взаємодіє з холінорецепторами постсинаптичноі мембрани м’язового волокна, деполяризує її. Так виникає потенціал дії м’язового волокна (ПДМВ). Таким чином, в процесі передачі збудження з нерва на м’яз можна виділити три ланки: електрична (нервовий імпульс), хімічна (утворення комплекса АХ-рецептор) і знову електрична (утворення ПДМВ). Для проведення збудження через нервово-м’язовий синапс необхідно 0,2-0,5 мс, через міжнейронний синапс – 1,5-2 мс. Цей час називається синаптичною затримкою. Він складається з часу, необхідного для вивільнення Ах з нервового закінчення, часу дифузії Ах від пресинаптичної мембрани до постсинаптичної, часу, необхідного для взаємодії Ах з рецепторами кінцевої пластинки. Запасів Ах в нервовому закінченні достатньо для проведення біля 10000 мото-нейронних імпульсів. При тривалій імпульсації мото-нейрона вміст Ах в кінцевих розгалуженнях може суттєво зменшитися, що призводить до розвитку пресинаптичного нервово-м’язового блоку. Комплекс «Ax-рецептор» швидко руйнується спеціальним ферментом – ацетилхолінестеразою (АхЕ) кінцевої пластинки. Це руйнування супроводжується відновленням вихідної проникності постсинаптичної мембрани, готовністю сприймати повторне збудження. Але при тривалій високочастотній імпульсації мотонейрона в синаптичній щілині може накопичуватися надлишок Ах, який не встигає руйнуватися. При цьому здатність постсинаптичної мембрани до генерації потенціалу кінцевої пластинки знижується і розвивається частковий чи повний постсинаптичний нервово-м’язовий блок. Обидва типи нервово-м’язового блоку (пре-і постсинаптичний) можуть бути причиною зниження скоротливої здатності м’язів (периферича втома).
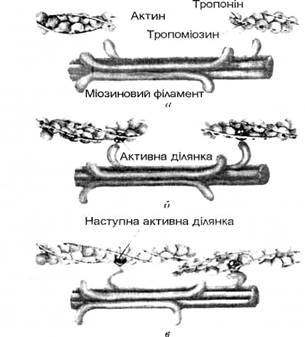
Мал.6. Будова нервово-м’язового синапса А – нервово-м’язовий синапс в розрізі Б – розприділення синапсів соматичного аксона в волокнах скелетного м’язу В -розприділення синапсів соматичного аксона в клітинах гладеньких м’язів. Механізм м’язового скорочення. Утворений під впливом нервових імпульсів ПДМВ розповсюджується на поверхні мембрани (по сарколемі) і по мембранах поперечних трубочок всередину волокна до цистерн поздовжніх трубочок. Електрична хвиля збудження деполяризуе мембрани цистерн і тим самим сприяє швидкому виходу іонів кальцію із цистерн у міжфібрилярний простір – до ниток актину та міозину. Цей вихід звільнених іонів кальцію знаменує собою початок скорочення м’яза, він розпочинається при деполяризації мембрани до 60 мВ і продовжується протягом всього часу деполяризаціі. Іони кальцію зв’язуються з тропоніном так, що молекула останнього деформується і штовхає тропоміозин у жолобок між двома ланцюгами актину в активне положення. В зв’язку з цим відкриваються раніше заблоковані тропоміозином ділянки актину для прикріплення поперечних містків міозину. Разом з тим іони кальцію активують АТФ-фазу міозинових головок, яка розщіплює АТФ з вивільненням енергії. Енергія АТФ забезпечує дію поперечних містків, які втягують тонкі актинові нитки в проміжки між товстими міозиновими нитками. Під впливом енергії АТФ в присутності іонів кальцію міозинові містки ритмічно прикріплюються до актинових ділянок актину і відкріплюються від них і таким чином тягнуть актинову нитку до середини саркомера, нагадуючи при цьому групу людей, які тягнуть довгий канат, перехоплюючи його руками. Рухаючись з двох сторін саркомера назустріч одна до другої, актинові нитки забезпечують вкорочення саркомера (від 2 мкм до 1,25 мкм); вкорочуються, скорочуючись, міофібрили, м’язові волокна і м’яз в цілому (мал.7)
Мал. 7. Механізм ковзання актинових і міозинових фі-ламентів (за Д.Уілмором і Д.Костіллом, 1997): м’язове волокно в стані розслаблення (а), в процесі скорочення (б) і повністю скорочене (в).
При надмірному розтягуванні м’яза або при сильному і швидкому його скороченні число взаємодіючих з актиновими нитками поперечних містків зменшується, їх зчеплення послаблюється, напруження м’яза знижується. Найбільше напруження м’яз розвиває при середній довжині саркомерів (2,0-2, 25мкм), коли зчеплення між нитками актину і поперечними містками міозину оптимальне. При сильному скороченні, коли актинові нитки в саркомері перекриваються між собою і міозинові нитки впираються в Z-перегородку, напруження м’яза зменшується. Іони кальцію затримуються біля міофіламентів (у міжфібрілярному просторі) всього декілька мілісекунд, а тоді внаслідок дії спеціальної кальцієвої помпи повертаються в цистерни. При цьому наступає розслаблення м’язового волокна. Тропонш і тропоміозин повертаються до виконання своєї гальмшної функції. Загальна схема послідовності процесів, які забезпечують м’язове скорочення, представлена на мал.8. Дата добавления: 2015-11-26 | Просмотры: 1780 | Нарушение авторских прав |