|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
ИНДУКЦИЯ ТОЛЕРАНТНОСТИ
Растворимые антигены. АПК обычно не способны к представлению растворимых антигенов, которые вследствие этого менее иммуногенны, но более толерогенны. Возможно, растворимые антигены блокируют рецепторы лимфоцитов и стимуляцию Т-клеток. Эмбриональный или неонатальныи период. Было установлено, что введение антигена в эти периоды жизни индуцирует толерантность. Возможно, в её основе заложены клональная элиминация и неспособность незрелых АПК представлять антиген. В-клетки эмбриона могут частично становиться толерантными, что связано с различиями в замещении их иммуноглобулиновых рецепторов (см. выше). Имеются доказательства, что а-фетопротеин как основной сывороточный белок эмбриона способен подавлять аутореактивные Т-лимфоциты. Оральный путь. Антигены, поступившие через кишечник, впервые обрабатываются макрофагами печени, которые удаляют иммуногенные агрегаты и другие комплексы, оставляя лишь растворимые толерогены. Кроме того, АПК в кишечнике могут быть специализированы для индукции толерантности, чтобы предотвращать иммунный ответ на пищевые антигены. АПК. Любое подавление функций АПК, включая введение антител к их молекулам ГКГС, создаёт условия для снижения иммуногенности и для индукции толерантности. Толерантность, индуцированная введением антител, может привести к усилению роста опухоли, трансплантата (эффект усиления). Действием антител к молекулам ГКГС объясняется, по-видимому, и благотворное влияние переливаний крови на выживаемость трансплантата почки. Антитела к CD4 молекуле частично эффективны в индукции Т-клеточной толерантности к антигену, вводимому в то же время. Высокие дозы антигена обычно более толерогенны, но повторные инъекции очень малых доз антигена тоже индуцируют Т-клеточную толерантность. Как правило, Т-клеточная толерантность индуцируется легче и на более длительный период, чем толерантность В-клеток. «Антигенное самоубийство». Конъюгирование антигена с токсическими препаратами, радиоизотопами и др. даёт возможность направленного разрушения специфических Т- и В-лимфоцитов без последствий для окружающих клеток. Подобный принцип применялся для элиминации опухолевых клеток с использованием конъюгатов специфических антител с токсинами (рис. 31). Комплексы антиген-антитело способны иногда индуцировать толерантность, блокируя рецепторы. В зависимости от природы и соотношения антител и антигена комплексы, напротив, могут быть высокоиммуногенными. Неиммуногенные носители, такие, как «свои» материалы или недеградируемые молекулы (например, D-аминокислоты), могут обеспечивать толерогенность гаптенов, которые на нормальных носителях являются антигенными. Это явление использовалось для подавления специфического IgE-ответа при аллергии. Аитиидиотипические антитела уже испытывались при лечении аутоиммунных заболеваний и могут оказаться полезными при трансплантации. К сожалению, супрессируя один идиотип, можно способствовать выработке других антител к тому же антигену, что ставит под сомнение целесообразность такого подхода. 22. ИДИОТИПЫ, АНТИИДИОТИПЫ И ИХ СЕТИ
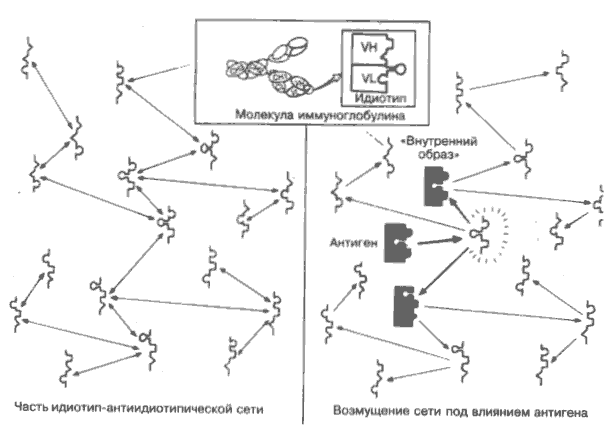
Учитывая, что антигенраспознающие участки антител Т-клеточных рецепторов кодируются в чрезвычайно большом разнообразии именно для того, чтобы распознавать практически любые предсказуемые молекулярные формы, нет ничего удивительного, что некоторые из них распознают друг друга. В таких случаях одну молекулу называют идиотипом (Id), а другую — антиидиотипом (анти-Id). Теоретически они могут быть не только антителами, но и молекулами Т-клеточных рецепторов (на рисунке вверху схематично показана локализация идиотипичес ких детерминант в активном центре антитела). Из этого можно сделать несколько важных выводов. Во-первых, очевидно, что это пример аутореактивности или аутоиммунитета. Во-вторых, неизвестно, как антиидиотип влияет на клетку, синтезирующую идиотипы, и наоборот, а также каковы последствия этих влияний на иммунный ответ в целом. Кроме того, сомнению подвергается привычное деление молекул на рецепторы и антигены. Действительно, молекула Ig-рецептора к определённому антигену сама оказывается антигеном для нескольких других Ig-молекул, для которых в свою очередь существуют рецепторы, и т.д. Таким образом, напрашивается вывод, что для каждого чужеродного антигена в этой практически бесконечной цепи где-то должна найтись молекула рецептора, по конфигурации точно соответствующая этому антигену, — его «внутренний образ». Иными словами, все антигены окружающей среды должны быть представлены в организме на лимфоцитах. В этом случае бактерии, вирусы и др. не вносят в организм ничего действительно чужеродного, и сеть из взаимодействующих между собой рецепторов (левая часть рисунка) должна существовать в организме и без их вторжения. Однако антигены из окружающей среды стимулируют синтез дополнительных антител, нарушающих равновесное состояние сети (правая часть рисунка), что приводит к образованию множества идиотипов и антиидиотипов, некоторые из которых (выделены на рисунке более темным цветом) частично или полностью соответствуют исходному антигену. В 1974 г. английский иммунолог Н. Ерне впервые сформулировал все эти возможности и объединил их в теорию идиотип-антиидиотипической сети, многочисленные аспекты которой нашли отражение во многих областях иммунологии. Сейчас трудно даже представить все возможные варианты практического приложения сетевой теории для регуляции иммунного ответа, лечения аутоиммунных заболеваний, создания вакцин и т.д. Имеется немного иммуннологических ситуаций, где бы не обсуждались вопросы, связанные с сетевой теорией. Дата добавления: 2014-12-12 | Просмотры: 1464 | Нарушение авторских прав |