|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
В ЛИМФОЦИТЫ (В-КЛЕТКИ)Ig, sIg — цитоплазматический и поверхностный иммуноглобулины соответственно, основная структура В-клетки, распознающая специфический антиген (рис. 12, 15). ГКГС II. Антигены, кодируемые в области ГКГС класса II, представлены главным образом на В-клетках и макрофагах и вовлекаются во взаимодействие с CD4 Т-клетками (рис. 13, 14, 17). CR2 — рецептор В-лимфоцитов к СЗ-компоненту комплемента; может быть вовлечён в генерацию анамнестического ответа. Декстраисульфат (ДС). Липополисахариды (ЛПС; например, эндотоксин сальмонеллы). Очищенное белковое производное туберкулина (PPD). Оказывают митогенное действие только на В-клетки, возможно, на разные стадии функционирования. Митоген лаконоса (МЛ). Оказывает митогенное действие на Т- и В-лимфоциты, если присутствуют оба типа клеток. К сожалению, идеального митогена для В-клеток человека не существует. Т-зависимость, Т-иезависимость. Синтез некоторых антител, главным образом IgM, происходит без помощи Т-хелперов и называется тимуснезависимым. В нём участвуют различные В-клетки, и, вероятно, в процессе эволюции он возник раньше других. В подавляющем большинстве случаев антителообразующая функция В-лимфоцитов является тимусзависимой. Апоптоз — запрограммированная гибель клеток. Процесс, в результате которого клетка гибнет без вреда для окружающих тканей. Очень большое количество В- и Т-клеток погибает по этому механизму, если в них не произошла перестройка генов рецепторов или если они становятся аутореактивными (рис. 36). 10. ЦЕНТРАЛЬНЫЕ ЛИМФОИДНЫЕ ОРГАНЫ И ЛИМФОПОЭЗ
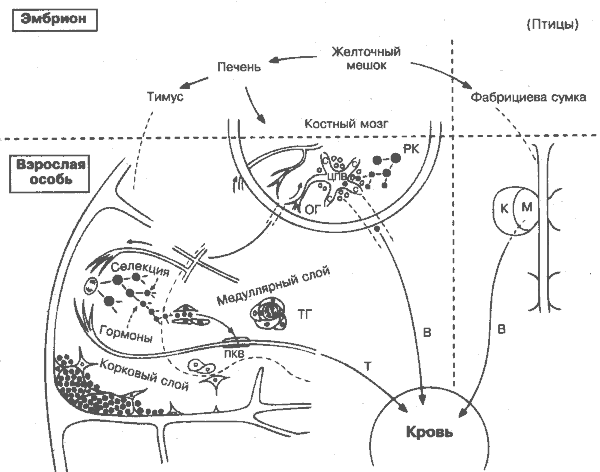
Первым серьёзным доказательством существования различающихся популяции лимфоцитов были взаимодополняющие последствия удаления у птиц в раннем возрасте тимуса (которое ведёт главным образом к нарушению клеточного иммунитета), и фабрициевой сумки (вызывающего нарушение антительного ответа). Поиск аналога фабрициевой сумки у млекопитающих продолжался долго. Одно время считалось, что это лимфоидная ткань, ассоциированная с кишечником. На самом деле аналога как такового не существует. В период внутриутробного развития В-лимфоциты созревают в печени, а у взрослых — в костном мозге Продукция В- и Т-лимфоцитов — весьма случайный и, на первый взгляд, расточительный процесс, который, в отличие от другой формы гемопоэза, включает перестройку генов, чтобы каждая клетка приобрела уникальный рецептор (рис. 14,15), и уничтожение всех тех клеток, у которых перестройка прошла неудачно и которые имеют рецепторы, способные распознавать молекулы «своего» и, таким образом, являются аутореактивными (рис. 36). Так как процесс распознавания антигена Т-клетками более сложен и задействует не только антиген, но и молекулы ГКГС (рис. 13, 14, 17). то и продукция Т-клеток — более усложнённый процесс, требующий двух раздельных этапов селекции: один для «своих» молекул ГКГС и другой против собственных антигенов. Вокруг самого тимуса споры не утихают до сих пор. Так и неясно, ограничено ли действие гормонов тимуса только границами органа, действительно ли тельца Гассаля — зона разрушения клеток. Зачем нужны два вида Т-клеточных рецепторов — a/b и g/d Как может один и тот же Т-клеточный рецептор подвергаться селекции дважды, причём в противоположных направлениях? Неизвестна также роль тимуса при старении. Здесь столько нерешённых вопросов, что остаётся удивляться, как это ещё не воскресили теорию Галена, согласно которой тимус — вместилище души! Желточный мешок — источник самой ранней гемопоэтической ткани, включая предшественники лимфоцитов. ФАБРИЦИЕВА СУМКА (бурса; лат. bursa Fabricii) - специальный орган для дифференцировки В-лимфоцитов у птиц — выступ заднего отдела клоаки, содержащий многочисленные фолликулы и крипты. Сумка достигает максимальных размеров через несколько недель после вылупления птенца, затем постепенно атрофируется. У млекопитающих аналогичный орган не найден, хотя его роль пытались приписывать аппендиксу, миндалинам и др. Медуллярый слои (М) — область, где первые стволовые клетки заселяют фолликулы бурсы. Корковый слой (К) — место пролиферации В-лимфоцитов. ПЕЧЕНЬ У млекопитающих печень — главный гемо- и лимфопоэтический орган в период внутриутробного развития. КОСТНЫЙ МОЗГ Родоначальные клетки (РК) для В-клеточной линии. Область гемопоэза (ОГ). Точная анатомическая локализация лимфопоэза в печени и костном мозге неизвестна. Предполагается, что он происходит наряду с гемопоэзом других клеток в тесном контакте с макрофагами и клетками стромы. Здесь же в результате неудачной перестройки генов иммуноглобулинов (рис. 9, 15) или чрезмерной аутореактивности погибает около 70% всех развившихся В-клеток. Синус (С) собирает дифференцированные клетки перед их выходом в кровь через центральную вену. ТИМУС Парный дольчатый орган, расположенный в верхнем отделе переднего средостения (у птиц — шеи); происходит из выпячивания в области 3-го и 4-го жаберных карманов. Аналогично бурсе максимальных размеров достигает в ранние сроки жизни, но атрофируется медленнее. Здесь костномозговые стволовые клетки преобразуются в зрелые Т-лимфоциты. Гормоны. Многочисленные растворимые факторы, экстрагированные из тимуса, стимулируют созревание Т-клеток, о чём свидетельствуют функциональные свойства, поверхностные маркёры или оба параметра. Общепринятой терминологии ещё нет, приводимый список далеко не полон: Тимозин a1 (м.м.3108), b1 (м. м. 8451), b4 (м. м. 4982). Тимопоэтин I, II (м. м.9562). Тимозин (м. м. 3108), В (м. м. 8451), В4 (м. м. 4982). Тимический гуморальный фактор (м. м. 3220). Тимостимулин (м. м. 12 000). Сывороточный фактор тимуса (тимулин) (м. м. 857). Корковый слой — интенсивно окрашиваемый внешний слой, упакованный лимфоцитами, разделёнными удлинёнными эпителиальными клетками. Основное место пролиферации и селекции Т-лимфоцитов. Медуллярный слой — внутренний, преимущественно эпителиальный, слой, куда перед выходом в кровь и лимфатические сосуды мигрируют лимфоциты из коркового слоя. Возможно, на границе медуллярного и коркового слоев проходят заключительные этапы селекции. Посткапиллярная венула (ПКВ), через которую лимфоциты поступают в вены тимуса и затем в кровь. Тельца Гассаля (ТГ) — специфичные для тимуса структуры, в которых эпителиальные клетки концентрически уплотнены и кератинизированы и где, возможно, удаляются клетки, подвергшиеся апоптозу. Селекция. Ввиду своей значимости и сложности процесс селекции Т-лимфоцитов вызвал пристальное внимание. На сегодня известно, что он состоит из следующих стадий:
11. ПЕРИФЕРИЧЕСКИЕ ЛИМФОИДНЫЕ ОРГАНЫ И РЕЦИРКУЛЯЦИЯ ЛИМФОЦИТОВ
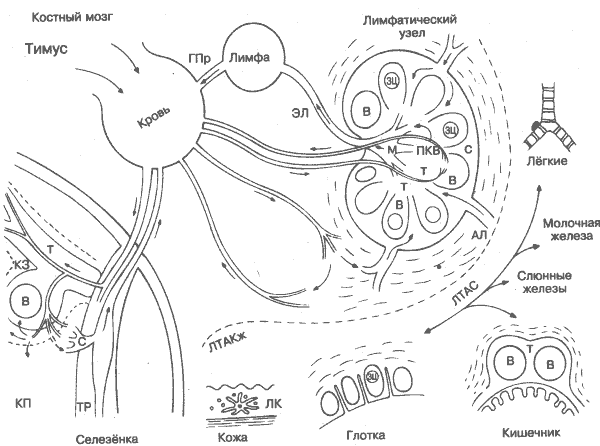
Способность к рециркуляции из крови в ткань и обратно через лимфоидную систему является уникальной для лимфоцитов и вместе с большой продолжительностью жизни и специфичностью к индивидуальным антигенам определяет их центральную роль в адаптивном иммунном ответе. В результате кругооборота лимфоцитов, особенно в селезёнке и лимфатических узлах, обеспечивается максимальный контакт антигенпредставляющих клеток, недавно получивших воздействие антигена, с Т- и В-лимфоцитами, преадаптированными к ответу на этот антиген. В других условиях это происходит крайне редко. Возросшие Т- и В-клеточные популяции распространяются по организму с готовностью к новой встрече с тем же антигеном, обеспечивая вторичный ответ. Разные типы лимфоцитов заселяют различные области лимфоидиых органов (Т- и В-зоны), что вызвано уникальностью местных хемотаксических факторов либо способностью различных лимфоцитов к распознаванию локальных структур, таких, как адгезивные молекулы на поверхности клеток сосудистого эндотелия или дендритных антигенпредставляющих клеток (рис. 7). В целом лимфатические узлы отвечают на антигены, вводимые в ткань, которые они дренируют, а селезёнка — на антигены, находящиеся в крови. Кишечник, лёгкие, молочные железы и внешние слизистые оболочки также имеют свои собственные, менее специализированные лимфоидные зоны, которые в некоторой степени имеют отдельную сеть рециркуляции клеток, обозначаемых как лимфоидная ткань, ассоциированная со слизистыми оболочками. Это относится и к коже — лимфоидная ткань, ассоциированная с кожей (ЛТАКж). В каждом случае локальная лимфоидная система специализирована в отношении антигена, который с наибольшей вероятностью поступает в эти зоны. Дата добавления: 2014-12-12 | Просмотры: 1433 | Нарушение авторских прав |