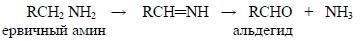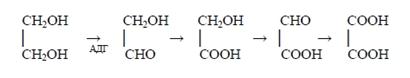|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Введение 4 страница
Рис. 7. Схема поступления ядовитых веществ через кожу
Следует учитывать, что соли многих металлов, соединяясь с жирными кислотами и кожным салом, могут превращаться в жирорастворимые соединения и проникать через барьерный слой эпидермиса (особенно ртуть и таллий). Механические повреждения кожи (ссадины, царапины, раны и пр.), термические и химические ожоги способствуют проникновению токсичных веществ в организм.
4.4. Транспорт токсичных веществ
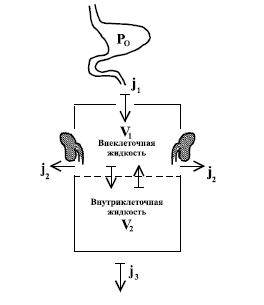
После поглощения любым путем вещества попадают в кровь, лимфу или какую-нибудь другую жидкость организма. Однако для большинства веществ наиболее важным средством транспортировки после всасывания до выделения является кровь. Токсичные вещества проникают в организм в виде молекул и ионов, однако, во внутренней среде они могут подвергаться гидролизу и полимеризации, образуя коллоидные частицы. В крови вещества находятся либо в свободном состоянии, либо связаны с каким-либо компонентом крови. Как и некоторые недиссоциированные молекулы, пары и газы могут физически растворяться в плазме в свободном виде. Различные токсичные вещества и их метаболиты транспортируются кровью в разных формах. Вещества могут быть связаны с эритроцитами или с компонентами плазмы. Выраженное сродство к эритроцитам имеют немногие вещества. Оксид углерода связывается с гемом, а мышьяк - с глобином гемоглобина. Свинец на 96 % переносится эритроцитами. Ртуть, содержащаяся в органических соединениях, и цезий также связываются с эритроцитами, а неорганическая ртуть - с альбумином плазмы крови. Большинство веществ проявляет сродство к белкам плазмы, преимущественно к альбуминам. Связь осуществляется ионными, водородными и ван-дер-ваальсовыми силами. Токсичные вещества могут образовывать комплексы с органическими кислотами плазмы или хелатные соединения с некоторыми ее компонентами. Как уже указывалось, из белков плазмы наиболее важным средством переноса является альбумин. Он имеет относительно крупную молекулу. При рН = 7,4 она имеет отрицательный заряд в 16 электронных единиц. Молекула содержит 100 отрицательных и 84 положительных группы (лиганды), так что может притягивать и переносить катионы и анионы. На поверхности молекул альбумина могут адсорбироваться нейтральные молекулы. Глобулины (а, в) могут связываться с небольшими молекулами различных веществ и ионами некоторых металлов (меди, цинка, железа), а также со всеми коллоидами. Фибриноген проявляет сродство только к очень маленьким молекулам. Белки плазмы могут переносить вещества, растворимые в липидах. Во многих случаях между белками плазмы и эритроцитами возникает конкуренция за различные вещества. Органические кислоты (молочная, глютаминовая, лимонная) образуют комплексы с различными веществами. К последним относятся щелочноземельные элементы, редкоземельные и некоторые тяжелые металлы, находящиеся в плазме в виде катионов. Обычно комплексы с органическими кислотами способны диффундировать и легко удаляются из тканей и органов. Присутствующие в крови природные вещества, вызывающие получение хелатных соединений, конкурируют с органическими кислотами за катионы, образуя стабильные хелаты. Путем хелатирования некоторые специальные белки (трансферрин, церулоплаз-мин) переносят ионы металлов (железа, меди). Органические лиганды веществ легко хелатируют двух- и трехвалентные ионы. Удаление токсичного вещества из крови зависит от его свойства связываться с компонентами крови. В некоторых случаях компоненты эритроцитов или плазмы могут удерживать яды продолжительное время. Таким образом, белки крови, способные связываться с токсичным веществом, помимо транспортной функции выполняют роль своеобразного защитного барьера. 4.5. Распределение и кумуляция Следующим этапом всасывания токсичного вещества в кровь является его распределение в организме. Одним из основных токсикологических показателей является объем распределения, т. е. характеристика пространства, в котором распределяется данное токсичное вещество. Существует три главных сектора распределения чужеродных веществ: внеклеточная жидкость (примерно 14 л для человека массой тела 70 кг), внутриклеточная жидкость (28 л) и жировая ткань, объем которой значительно варьируется. Объем распределения зависит от трех основных физико-химических свойств данного вещества: водорастворимости, жи-рорастворимости и способности к диссоциации. Водорастворимые соединения способны распространяться во всем водном секторе организма (около 42 л), жирорастворимые вещества депонируются преимущественно в липидах. Основным препятствием для распространения водорастворимых веществ являются плазменные мембраны клеток. Именно процесс диффузии через этот барьер будет определять накопление вещества внутри клеточного объема, т. е. переход от распределения в 14 л воды (внеклеточная жидкость) к распределению в 42 л. Для анализа распределения чужеродного вещества в организме достаточно рассмотреть двухкамерную модель. Эта максимально упрощенная модель позволяет понять, как меняются концентрации токсичных веществ в клеточном и внеклеточном секторах организма (рис. 8). Камера V1 включает всю внеклеточную жидкость с концентрацией токсичного вещества С, что соответствует уровню препарата в плазме крови. Камера V2 содержит внутриклеточную жидкость с концентрацией токсичного вещества С, где k - коэффициент пропорциональности. Этот коэффициент условно определяет сродство ткани к данному веществу. В количественном отношении это сродство может варьировать в очень широких пределах.
Рис. 8. Двухкамерная модель распределения ядов в организме
Введение коэффициента k для определения концентрации в клеточном секторе является первым приближением процесса разведения вещества, поступающего в кровоток. Оно применимо в тех случаях, когда процессы поступления или элиминации проходят с постоянными времени, на порядок большими, чем время полной циркуляции крови. Принято считать, что в каждый момент имеется равновесное распределение вещества в организме. Это приближение достаточно для клинических целей. Нарушение условия равновесия приводит к усложнению модели. Процесс неравномерного распределения токсичных веществ в организме, связанный с их накоплением в отдельных структурах, делает понятие объема распределения в кинетической модели условным. Поэтому под этим термином часто понимают не истинный объем соответствующего отдела организма, а некий коэффициент пропорциональности, связывающий общую дозу вещества (Ро), введенного в организм, и его концентрацию (С), определяемую в плазме: V = Ро / С. Наиболее точно объем распределения можно вычислить при разовом внутривенном введении вещества, так как в этом случае известно количество вещества, поступившего в кровь. Если расчетный объем распределения превышает количество внеклеточной жидкости, то следует думать о частичном проникновении вещества в клетки. В случае если объем распределения будет больше, чем объем всей жидкости организма, то это означает, что коэффициент связывания препарата тканями больше единицы и происходит его внутриклеточное накопление. Итак, судьба вещества, поступающего в организм из желудочно-кишечного тракта и распределяющегося в двухкамерной системе, может быть представлена в виде направленных потоков: где j1 - поток вещества, всасывающегося из желудка; j2 - поток экскреции; j3 - условный поток утилизации препарата в тканях (метаболическое превращение); С - концентрация вещества в плазме; к - коэффициент связи вещества с белками сектора V2. По распределению в тканях и проникновению в клетки химические вещества можно разделить на две основные группы: неэлектролиты и электролиты. Неэлектролиты, растворяющиеся в жирах и липидах, подчиняются закону Овертона и Майера, согласно которому вещество тем скорее и тем в большем количестве проникает в клетку, чем больше его растворимость в жирах, иначе говоря, чем больше коэффициент распределения (К) между жирами и водой: К = растворимость в масле / растворимость в воде. Это объясняется тем, что оболочка клеток содержит много липидов. Для данной группы химических веществ барьеров в организме не существует: распределение неэлектролитов в организме при динамическом поступлении определяется в основном условиями кровоснабжения органов и тканей. Это подтверждается следующими примерами. Мозг, содержащий много липидов и имеющий богатую кровеносную систему, насыщается этиловым эфиром очень быстро, в то время как другие ткани, содержащие много жира, но с плохим кровоснабжением, насыщаются эфиром очень медленно. Аналогично происходит насыщение анилином. Удаление неэлектролитов из тканей также зависит в основном от кровоснабжения: после прекращения поступления яда в организм быстрее всего освобождаются от него органы и ткани, богатые кровеносными сосудами. Из мозга, например, удаление анилина происходит значительно быстрее, чем из околопочечного жира. В конечном же итоге, неэлектролита после прекращения поступления их в организм распределяются во всех тканях равномерно. Способность электролитов проникать в клетку резко ограничена и, как полагают, зависит от заряда ее поверхностного слоя. Если поверхность клетки заряжена отрицательно, она не пропускает анионов, а при положительном заряде она не пропускает катионов. Распределение электролитов в тканях очень неравномерно. К особенностям распределения электролитов в организме относится прежде всего их способность быстро удаляться из крови и, накапливаясь в отдельных органах, образовывать в организме депо. Время задержки определяется сродством вещества или его метаболита к зоне связывания. Время, необходимое для выведения 50 % вещества из депо, называется периодом полувыведения. Если скорость абсорбции выше скорости выведения, токсичное вещество будет накапливаться. Распределение вещества -не статический, а динамический процесс. Со временем токсичное вещество может распределяться в различные камеры в результате обмена веществ или физико-химических изменений среды (рН, степени диссоциации). Вещества, которые находятся в крови в виде одновалентных катионов (литий, цезий, рубидий) или в виде анионов с валентностями от 1 до 6 (хлор, бром, полоний), легко диффундируют в жидкостях в организме и распределяются равномерно во всех органах и тканях. Отложение в ретикуло-эндотелиальной системе органов. Часть неподвижных или блуждающих (фагоциты) клеток органов могут находить, захватывать и уничтожать чужеродные тела (коллоиды, частицы, микроорганизмы). Эти клетки представляют собой ретикуло-эндотелиальную систему органов и тканей. Интерес представляет задержка коллоидов. С увеличением размера коллоидных частиц они накапливаются преимущественно в печени. С уменьшением размера частиц достигается более равномерное их распределение по другим органам. Отложение в жировых тканях. Жирорастворимые вещества проявляют большое сродство к тканям и органам, богатым липидами и жирами: жировой ткани, эндокринным железам, нервным волокнам. Многие из этих веществ (ДДТ) могут накапливаться в жировой ткани. Некоторые нейротоксичные вещества оказывают свое действие благодаря тому, что растворяются в липидах миелиновой оболочки нервных волокон. Отложение в костной ткани. Многие вещества проявляют особое сродство к костной ткани: легкие и щелочноземельные элементы, некоторые коллоиды. Минеральная часть кости состоит из минералов гидроксиапатита Са10(РО4)6(ОН)2. Остео тропные вещества могут откладываться в минеральных компонентах кости с помощью двух основных механизмов: а) ионообмена катионов Са2+ из гидроксиапатита либо анионов с фосфатными анионами или анионами гидроксильной группы; б) абсорбции коллоидов на поверхности костных кристаллов. У костных кристаллов огромная поверхность (100 м2/г), на которой может адсорбироваться один или множество слоев коллоидных частиц, которые затем покрываются следующим минеральным слоем. Отложение в волосах и ногтях. Некоторые тяжелые металлы (свинец, цинк, кадмий, ртуть) проявляют сродство к SH-группам кератина, находящегося в волосах.
4.6. Биотрансформация токсичных веществ
Большинство чужеродных органических, а также некоторые неорганические вещества, претерпевают в организме метаболические превращения благодаря катализу внутри- и внеклеточными ферментами. Эти реакции обычно приводят к образованию производных, молекулы которых более полярны, чем у исходных веществ, поэтому они легче выводятся из организма. Все ткани, в том числе клетки печени, почек, кишечника и плаценты, обладают способностью (различной у разных органов) к метаболизму посторонних веществ, но основным местом биотрансформации является печень, и, в частности, клетки паренхимы. Биотрансформация катализируется в соответствии с химической структурой чужеродного вещества ферментами, находящимися в различных компонентах клетки (растворимая фракция цитоплазмы, эндоплазматическая сеть, митохондрии, лизосомы, ядро и т. д.). Многие реакции катализируются ферментами гладкой эндоплазматической сети (микросомы). При биотрансформации, претерпеваемой чужеродными веществами в организме, могут быть выделены реакции двух типов: 1) реакции фазы I, в основном окисление, восстановление и гидролиз; 2) реакции фазы II, представляющие собой биосинтетические реакции конденсации, с помощью которых чужеродные вещества или их метаболиты, образующиеся по реакциям фазы I, соединяются с эндогенными субстратами. Сами реакции фазы I могут быть, в свою очередь, разделены на два подкласса: катализируемые микросомными ферментами (ферментами эндоплазматической сети) и немикросомными ферментами.
4.6.1. Реакции окисления, катализируемые микросомными ферментами (микросомальное окисление)
Окисление может быть схематически представлено следующим образом:
Для реакции нужны молекулярный кислород и электроны. Один из атомов молекулы О2 включается в чужеродное вещество, а другой восстанавливается с образованием молекулы воды. По этой причине а ферменты, катализирующие эти реакции, известны как монооксигеназы (МО) или оксидазы смешанной функции. Детальный механизм этой реакции до сих пор не выяснен полностью, но установлено, что гемопротеид, ингибированный оксидом углерода и известный под названием цитохром Р-450, играет центральную роль при окислении. Простейший детокси-цирующий цикл заключается в следующем (рис. 9). Попавшие в организм экзогенные чужеродные вещества (RH) соединяются с альбумином (А) и в виде комплекса (RHA) транспортируются в печень. Часть из них может попадать в печень в свободном виде. Здесь на цитохроме Р-450 в мембранах эндоплазматической сети гепатоцита происходит окисление ксенобиотика. Цитохром Р-450 - это сложный белок, состоящий из двух частей: апофермента - собственно белковой части и про-стетической группы - гема. Апофермент выполняет регулятор-ную функцию и может связывать сотни самых различных соединений. Гем обладает способностью переводить молекулярный кислород из неактивной формы и использовать его в реакциях окисления, которых насчитывается несколько десятков. Он работает в составе окислительно-восстановительной цепи, поставляющей необходимые для активации кислорода электроны. В качестве поставщика последних выступает метаболит гликогена - восстановленный никотинамидаденинди-нуклеотидфосфат (НАДФН). Концентрация цитохрома Р-450 варьирует от одной ткани к другой и обычно выше всего в печени, что объясняет, по крайней мере частично, высокую метаболическую способность печени по отношению к большинству чужеродных веществ. Сам цитохром может быть ингибирован различными чужеродными веществами с образованием денатурированной формы цитохрома Р-420, не способного к участию в окислении.
Рис. 9. Схема механизма окисления гидрофобных веществ цитохромом Р-450: 1Ш - гидрофобное вещество; А - альбумин; ЯОН - окисленное гидрофобное вещество
Примеры микросомальных реакций окисления: - гидроксилирование ароматических соединений (например, бензола в фенол) с образованием промежуточного метаболита - эпоксибензола:
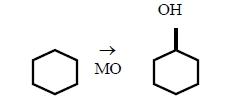
- гидроксилирование алифатических заместителей в ароматических соединениях, например, н-пропилбензола в этилфенил-карбинол. - гидроксилирование алициклических соединений, например, образование циклогексанола из циклогексана.
- окислительное О-дезалкилирование, например, 4-нитроанизола в 4-нитрофенол.
- окислительное N-дезалкилирование, например, деметили-рование диметилформамида в монометилформамид.
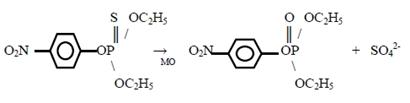
- S-дезалкилирование, например, деметилирование тиоэфиров - окислительное десульфирование, например, превращение паратиона в параоксон
4.6.2. Окисление, катализируемое немикросомными ферментами (немикросомальное окисление) Окисление может катализироваться ферментами, присутствующими в митохондриях, растворимой фракции цитоплазмы или в плазме. Примером может служить окисление первичных аминов в альдегиды аминоксидазой:
К этому же классу реакций биотрансформации относится окисление этанола и других спиртов алкогольдегидрогеназой (АДГ) в присутствии НАД (никотинамидадениндинуклеотид). Указанные реакции являются процессами, в результате которых малотоксичное вещество превращается в более токсичное, чем исходное. Такое явление называется летальным синтезом. Яркий пример такого рода превращения - метаболизм метилового спирта, токсичность которого полностью определяется продуктами его окисления - формальдегидом и муравьиной кислотой:
Метаболизм этилового спирта начинается с образования ацетальдегида, который по крайней мере на порядок токсичней исходного продукта:
Тяжесть отравления этиленгликолем прямо пропорциональна степени окисления его до щавелевой кислоты:
4.6.3. Реакции восстановления, катализируемые микросомными ферментами (микросомальное восстановление)
В микросомальной фракции гепатоцитов содержатся ферменты, не только окисляющие, но и чужеродные восстанавливающие органические соединения. Ароматические нитросоединения восстанавливаются в амины с промежуточным образованием гидроксиаминов с помощью микросомных ферментов в присутствии НАДФ в отсутствие кислорода. Нитробензол восстанавливается таким образом в анилин:
Микросомные азоредуктазы катализируют восстановление азопроизводных в амины. Например, n-диметиламинбензол восстанавливается в диме-тилфенилендиамин и анилин:
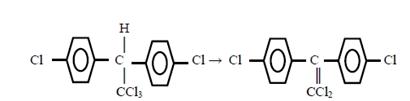
Микросомные ферменты могут также дегидрогалогениро-вать посторонние вещества по восстановительному механизму, например, ДДТ превращается в ДДЭ:
4.6.4. Немикросомальное восстановление
Реакции восстановления, катализируемые немикросомными ферментами, включают: - восстановление дисульфидов в меркаптаны; - восстановление гидроксамовых кислот в амиды; - восстановление N-оксидов в амины; - дегидроксилирование ароматических или алифатических гидроксипроизводных.
4.6.5. Гидролиз, катализируемый микросомными и немикросомными ферментами
Эстеразы и амидазы, присутствующие в различных компонентах клетки и в плазме, катализируют гидролиз многих сложных эфиров и аминов. Алифатические нитрилы могут гидроли-зоваться с образованием цианид-иона С№:
4.6.6. Конденсация
Конденсация чужеродных веществ или их метаболитов с различными эндогенными субстратами обычно дает более полярные производные, которые легко выводятся из организма с мочой или желчью. Основными реакциями являются следующие: 1. Конденсация с глюкуроновой кислотой. Как и большинство реакций конденсации, эта реакция протекает в две фазы. Сначала синтезируется донор глюкуроновой кислоты (уридиндифосфат-а-Б-глюкозидуроновая кислота). Затем следует связывание глюкуроновой кислоты с чужеродным веществом или его метаболитом. Синтез донора катализируется ферментами растворимой фракции цитоплазмы, а связывание глюкуроновой кислоты - микросомными ферментами. Глюкуроновые конденсаты обычно классифицируют следующим образом: а) О-глюкурониды, образованные из фенольных производных, спиртов, карбоновых кислот и гидроксиламинов; б) N-глюкурониды, образованные из ароматических производных; в) S-глюкурониды, образованные из меркаптановых производных. 2. Сульфоконденсация. Донор - аденозил-з-фосфат-5-фосфосульфат, который передает свою сульфогруппу с помощью сульфотрансферазы фенольному производному, спирту или амину. 3. Метилирование. Метильная группа передается от з-аденозилметио-нина аминопроизводному фенола или меркапто-группе. В метильные производные могут быть превращены также некоторые неорганические вещества, например, Se. 4. Ацетилирование. В этой реакции посредником является конфермент А. В процесс вовлекаются главным образом ароматические амины, сульфонамиды и производные гидразина. 5. Конденсация с глицином. Ароматические кислоты, например, бензойная, при конденсации с глицином образуют гип-пуровую кислоту или ее гомологи. Посредники в данной реакции АТФ (аденозинтрифосфат) и конфермент А. 6. Конденсация с глутатионом. Различные ароматические соединения, например, бензол, нафталин, полициклические углеводороды, образуют с глутатионом премеркаптуровые кислоты, которые представляют собой продукты взаимодействия с L-ацетилцистеиновой фракцией глутатиона; 7. Конденсация с серой. Эта реакция квазиселективно затрагивает цианидную группу, которая соединяется с серой тиосульфата с образованием тиоцианата: 4.6.7. Различные биопревращения Существуют также метаболические превращения, которые не могут быть отнесены ни к одному из вышеуказанных классов, например, раскрытие колец гетероциклических соединений или, наоборот, циклизация каких-нибудь веществ. Одни и те же химические соединения могут претерпевать различные метаболические превращения - как параллельно, так и последовательно. Пути превращения чужеродных веществ в организме сильно не схожи у различных лиц, а с течением времени изменяются и у одного и того же индивидуума. На метаболическую способность организма влияют генетические, физиопатологические, а также различные экзогенные факторы. К сожалению, сведения о метаболизме большого количества соединений недостаточны. Пути метаболизма токсичных веществ приходится изучать в основном на животных. Сложная природа видовых различий в метаболизме чрезвычайно затрудняет интерпретацию экспериментальных результатов, а возможность их использования для оценки метаболизма у человека очень ограничена. 4.7. Пути выведения чужеродных веществ из организма Пути и способы естественного выведения чужеродных соединений из организма различны. По их практическому значению они располагаются следующим образом: почки - кишечник -легкие - кожа. Выделение токсичных веществ через почки происходит с помощью двух основных механизмов - пассивной диффузии и активного транспорта. В результате пассивной фильтрации в почечных клубочках образуется ультрафильтрат, который содержит многие токсичные вещества, в том числе неэлектролиты, в той же концентрации, что и в плазме. Весь нефрон можно рассматривать как длинную полупроницаемую трубку, через стенки которой происходит диффузный обмен между протекающей кровью и формирующейся мочой. Одновременно с конвективным потоком вдоль нефрона токсичные вещества диффундируют, подчиняясь закону Фика, через стенку нефрона обратно в кровь (так как внутри нефрона их концентрация в 3-4 раза выше, чем в плазме) по градиенту концентрации. Количество вещества, которое покинет организм с мочой, зависит от интенсивности обратной реасорбции. Если проницаемость стенки нефрона для данного вещества высокая, то на выходе концентрации в моче и в крови выровняются. Это означает, что скорость выведения будет прямо пропорциональна скорости мочеобразования, а количество выводимого вещества будет равно произведению концентрации свободной формы яда в плазме на скорость диуреза l = kvm. Это минимальное значение выведенного вещества. Если стенка почечного канальца полностью непроницаема для токсичного вещества, то количество выделяемого вещества максимально, не зависит от скорости диуреза и равно произведению объема фильтрации на концентрацию свободной формы токсичного вещества в плазме: l = kvф. Реальное выведение ближе к минимальным значениям, чем к максимальным. Проницаемость стенки почечного канальца для водорастворимых электролитов определяется механизмами «неионной диффузии», т. е. пропорциональна, во-первых, концентрации недиссоциированной формы; во-вторых, степени растворимости вещества в липидах. Эти два обстоятельства позволяют не только прогнозировать эффективность почечной экскреции, но и управлять, хотя и ограниченно, процессом реасорбции. В почечных канальцах неэлектролиты, хорошо растворимые в жирах, путем пассивной диффузии могут проникать в двух направлениях: из канальцев в кровь и из крови в канальцы. Определяющим фактором почечного выделения является концентрационный индекс (К): К = Св моче/ Св плазме, где С - концентрация токсического вещества. Значение К < 1 свидетельствует о преимущественной диффузии веществ из плазмы в мочу, при значении К > 1 - наоборот. Направление пассивной канальцевой диффузии ионизированных органических электролитов зависит от рН мочи: если канальце-вая моча более щелочная, чем плазма, в мочу легко проникают слабые органические кислоты; если реакция мочи более кислая, в нее проходят слабые органические основания. Кроме того, в почечных канальцах осуществляется активный транспорт сильных органических кислот и оснований эндогенного происхождения (например, мочевой кислоты, холина, гистамина и пр.), а также чужеродных соединений сходной с ними структуры с участием тех же переносчиков (например, чужеродных соединений, содержащих аминогруппу). Металлы выделяются преимущественно почками не только в свободном состоянии, если они циркулируют в виде ионов, но и в связанном, в виде органических комплексов, которые подвергаются клубочковой ультрафильтрации, а затем через канальцы проходят путем активного транспорта. Выделение токсичных веществ, поступивших перорально, начинается уже в полости рта, где в слюне обнаруживаются многие электролиты, тяжелые металлы и т. д. Большинство металлов, задерживающихся в печени, может связываться с желчными кислотами (марганец) и с желчью выделяться через кишечник. Таким образом, удалению через кишечник с калом подвергаются вещества: 1) не всосавшиеся в кровь при их пероральном поступлении; 2) выделенные с желчью из печени; 3) поступившие в кишечник через мембраны его стенки. В последнем случае основным способом транспорта ядов служит пассивная их диффузия по градиенту концентрации. Большинство летучих неэлектролитов выделяется из организма в основном в неизменном виде с выдыхаемым воздухом. Начальная скорость выделения через легкие газов и паров определяется их физико-химическими свойствами: чем меньше коэффициент растворимости в воде, тем быстрее происходит их выделение, особенно той части, которая находится в циркулирующей крови. Выделение их фракции, депонированной в жировой ткани, задерживается и происходит гораздо медленнее, тем более, что это количество может быть очень значительным, так как жировая ткань может составить более 20 % от общей массы человека. Например, около 50 % поступившего ингаляционным путем хлороформа выделяется в течение первых 8-12 ч, а остальная часть - во второй фазе выделения, которая продолжается несколько суток. Многие неэлектролиты, подвергаясь медленной биотрансформации в организме, выделяются в виде основных продуктов распада: воды и углекислоты, которая выходит с выдыхаемым воздухом. Через кожу, в частности с потом, выходят из организма многие вещества - неэлектролиты, а именно: этиловый спирт, ацетон, фенолы, хлорированные углеводороды и пр. Однако, за редким исключением (например, концентрация сероуглерода в поте в несколько раз выше, чем в моче), общее количество удаляемого таким образом токсичного вещества невелико и не играет существенной роли. Дата добавления: 2015-02-05 | Просмотры: 790 | Нарушение авторских прав |