|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
КЛЕТОЧНОЕ ЯДРО
Клеточные ядра образуются только из ядер. Главные функции клеточного ядра: 1) хранение наследственной информации; 2) передача наследственной информации в цитоплазму с помощью транскрипции, т. е. синтеза переносящей информацию mРНК и передача информации дочерним клеткам при репликации – делении клеток и ядер. Ядро чаще всего расположено в центре клетки, и только у растительных клеток с центральной вакуолью – в пристеночной протоплазме. Оно может быть сферическим, яйцевидным, чечевицеобразным, реже сегментированным, вытянутым в длину или веретеновидным, но обычно повторяет форму клетки. Диаметр ядра варьирует в пределах от 0,5 мкм (у грибов) до 500 мкм (в некоторых яйцеклетках), в большинстве случаев он меньше 5мкм. Положение, форма и размеры ядра могут изменяться, часто параллельно с изменениями интенсивности метаболизма. Ядро состоит из нуклеоплазмы, хромосом (хроматина), ядрышек и ядерной оболочки (представляющей собой часть эндоплазматического ретикулума).
3.5.1 Нуклеоплазма Основная масса клеточного ядра – нуклеоплазма – содержит жидкую часть, ядерный матрикс (нечто вроде опорной сети) и различные включения. Жидкая часть сходна по составу с соответствующим компонентом цитоплазмы – здесь тоже содержатся ферменты и промежуточные продукты метаболизма, в частности гликолиза. Ядерный матрикс представляет собой с трудом выявляемый трехмерный «каркас», который состоит из кислых белков и пронизывает всю нуклеоплазму.
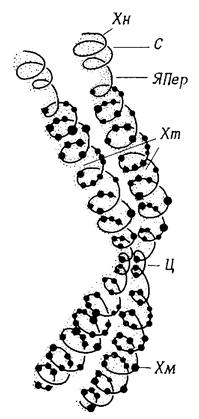
3.5.2 Хромосомы Хромосомы – это вытянутые в длину нуклеопротеидные структуры. Они удваиваются в результате идентичной репродукции перед каждым клеточным делением, а затем распределяются поровну между дочерними клетками. Поэтому каждая отдельная хромосома встречается во всех клетках данного индивидуума в одной и той же форме и несет идентичную информацию. На протяжении клеточного цикла происходит лишь смена двух физиологических форм: 1) транспортной (во время деления ядер; хромосомы компактные, палочковидные или колбасовидные, ясно различимые); 2) функциональной (в промежутках между делениями; хромосомы разрыхленные, нитевидные, длинные и неразличимы по отдельности). Хромосомы состоят из хроматина, который содержит около 40% ДНК, 40% гистонов, почти 20% негистоновых хромосомных белков и немного РНК. Он специфически окрашивается (отсюда и названия хромосома, хроматин) реактивами, выявляющими ДНК (например, реактивом Фёльгена) в красно-фиолетовый цвет. Гистоны – это хромосомные белки (5 видов) основного характера с высоким содержанием аргинина и лизина. Негистоновые хромосомные белки – главным образом кислые белки. Существует больше ста, вероятно несколько сотен, таких белков. К ним относятся белки, ответственные за движение хромосом (актин, миозин, тубулин), ферменты для синтеза РНК и ДНК (полимеразы) и для специфической модификации гистонов и других хромосомных белков, а также, вероятно, белки, регулирующие активность отдельных генов. Хромосомы во время деления ядер имеют длину 0,2–20 мкм и вначале состоят из двух лежащих рядом идентичных хроматид, которые потом отделяются друг от друга, причем каждая становится одной из дочерних хромосом. Главный элемент каждой хроматиды – нуклеопротеидная структура, которая помимо белков содержит единственную, очень длинную двойную спираль ДНК. Эта структура имеет вид закрученной толстой (15–25 нм) нити – хромонемы, на которой расположены многочисленные округлые (вследствие плотной упаковки ДНК) сильно красящиеся и хорошо видимые хромомеры (рис. 3.5). В месте первичной перетяжки хромосомы – центромере – обе хроматиды прочно связаны и в определенных фазах ядерного деления согнуты. Во время деления ядра центромера становится местом прикрепления нитей веретена, приводящих хроматиды в движение. У некоторых хромосом имеется еще ядрышковая перетяжка (SAT-зона). В этом месте хромосома имеет толщину всего лишь 7 нм и мало закручена, поэтому SAT-зоны не окрашиваются (SAT означает sine acido thymonucleinico, т. е. «без ДНК»). Отделяемый такой зоной короткий участок хромосомы называют сателлитом, а всю хромосому – SAT-хромосомой. Тесно примыкая к ядрышковой перетяжке, часто напротив сателлита, находится организатор ядрышка; это та часть нуклеопротеидной структуры, которая образует ядрышко после разделения ядра (рис. 3.5).
Рис. 3.5 – Клеточное ядро и хромосомы. А. Ядро во время интерфазы. ГХ – гетерохроматин; Риб – рибосомы; РНП – рибонуклеопротеидные частицы; ЭР – эндоплазматический ретикулум; ЭХ – эухроматин; Я– ядрышко; ЯО – ядерная оболочка; ЯП– ядерные поры; ЯТ– ядерные тельца. Б. SAT-хромосома во время деления ядра. С – сателлит; Хм – хромомеры; Хн – хромонема; Хт – хроматиды; Ц – центромера; ЯПер – ядрышковая перетяжка. В. Хромоцентрическое ядро (по Heitz). Хц – хромоцентр; Я – ядрышко
Между делениями ядер – в интерфазе – чаще всего нельзя различить отдельных хромосом. Разрыхленный, волокнистый хроматин распределен по всему объему ядра. Разрыхление структуры хромосом – необходимое условие для транскрипции, т. е. передачи информации, содержащейся в ДНК, путем образования mРНК. Уплотненный хроматин в отношении транскрипции неактивен, тогда как разрыхленный может быть неактивным или активным в зависимости от процессов регуляции. Сильно окрашивающиеся участки хроматина называют гетерохроматином в отличие от эухроматина, который после деления ядра разрыхляется. Гетерохроматин в отношении транскрипции неактивен и в отношении репликации ДНК ведет себя иначе, чем эухроматин. Факультативный гетерохроматин бывает гетерохроматичным только временами. Конститутивный гетерохроматин всегда гетерохроматичен. Он состоит из многократно повторяющихся последовательностей оснований, не информативен (не содержит генов) и поэтому всегда неактивен в отношении транскрипции. Соматические клетки имеют меньше конститутивного гетерохроматина, чем половые клетки, так как во время эмбрионального развития гетерохроматичекие участки вырезаются из ДНК ферментами. По всей длине каждой хромосомы (или, после репликации каждой хроматиды) проходит непрерывная двойная спираль ДНК, которая у высших организмов состоит более чем из 108 пар оснований. Гены линейно распределены вдоль этой двойной спирали и составляют вместе до 25% ДНК. Ген – это функциональная единица ДНК, содержащая информацию для синтеза полипептида или РНК (rРНК, tPHK). Средняя длина гена около 1000 пар оснований, что составляет 340 нм вытянутой двойной спирали ДНК. Последовательность, оснований в каждом гене уникальна. Между генами находятся спейсеры – неинформативные отрезки ДНК различной длины (иногда более 20 000 пар оснований), которые, по-видимому, имеют значение для регулирования транскрипции соседнего гена. Возможно, что функциональная единица «спейсер + ген» соответствует цитологической единице «отрезок хромонемы + 1 хромомер». Однако хромомеры могут содержать и несколько генов. Избыточные гены представлены большим числом (до 104 и более) идентичных копий; таковы, например, гены для tPHK > rРНК, 5S-PHK и гистонов, а также для продуктов, синтезируемых в больших количествах. Повторяющиеся последовательности – это последовательности нуклеотидов, многократно представленные в ДНК. Умеренно повторяющиеся последовательности – более длинные последовательности длиной в среднем 300 пар нуклеотидов с 102–104 повторениями. К ним относятся избыточные гены, а также большинство спейсеров. Высокоповторяющиеся последовательности с 105–106 повторениями образуют конститутивный гетерохроматин), они всегда неинформативны. Около 75% всего хроматина не участвует в транскрипции: это высокоповторяющиеся последовательности и нетранскрибируемые спейсеры. Весь фонд генетической информации каждого клеточного ядра – геном–распределен между некоторым постоянным числом хромосом. Это число, n, специфично для данного вида (или подвида). У лошадиной аскариды Parascaris equorum var. univalens оно равно 1, у кукурузы 10, у человека 23, у водоросли Netrium digitus около 600. Хромосомы одного набора различаются по величине, картине хромомер, положению перетяжек и содержанию информации. Гаплоидные клетки содержат один набор хромосом (n), а диплоидные – два (2 n, так что вся информация представлена дважды). В полиплоидных клетках несколько наборов хромосом (4n, 8n, 16n и т.д.). Половые клетки гаплоидны. У высших растений и животных соматические клетки диплоидны и содержат один отцовский и один материнский набор хромосом. Гаплоидные клетки образуются из диплоидных в результате мейоза, а диплоидные – из гаплоидных в результате оплодотворения. Полиплоидные клетки возникают из диплоидных путем эндомитоза – преждевременно прерванного деления ядра: после полной репликации и разделения хроматид дочерние хромосомы остаются в одном клеточном ядре, вместо того чтобы распределиться между двумя ядрами. Этот процесс может повторяться многократно. Аномалии при образовании половых клеток могут приводить к полиплоидизации всего организма. Хромосомы диплоидного ядра могут быть сгруппированы попарно, по две гомологичные хромосомы. Большинство из них (так называемые аутосомы) попарно идентичны. Только две половые хромосомы (гетерохромосомы), определяющие пол особи, у самцов неодинаковы: это хромосомы X и Y; большую часть последней занимает конститутивный гетерохроматин; у самок – две Х-хромосомы (у бабочек, птиц и ряда других животных наоборот: самцы имеют XX, самки – XY).
3.5.3 Ядрышко Ядрышки – это округлые, особенно уплотненные участки клеточного ядра диаметром обычно меньше 1 мкм. Ядра диплоидных клеток содержат 1–7, в среднем 2 ядрышка, а иногда (например, ядра у дрожжей, в сперматоцитах, микронуклеусы инфузорий) не имеют их совсем. Ядрышки осуществляют синтез рибосомальной РНК. В соответствии с этим главной составной частью ядрышка является ядрышковая ДНК, которая принадлежит организатору ядрышек одной из SAT-хромосом. Ядрышки содержат более 80% белка и около 15% РНК. В электронном микроскопе можно различить: 1) ядрышковый хроматин; 2) рибонуклеопротеидные (РНП-) фибриллы диаметром 5–10 нм и длиной 20–40 нм. Это ранние промежуточные продукты в процессе образования rРНК из пре-rPHK; 3) РНП-гранулы диаметром 15–20 нм – более поздние промежуточные продукты; 4) основную массу из белков и РНК, которую пронизывает сеть ядерного матрикса; 5) мелкие вакуоли; 6) гетерохроматин, связанный с ядрышком, который прилегает к ядрышку снаружи и проникает в него. С помощью светового микроскопа в некоторых ядрышках можно видеть нуклеолонему. Во время деления ядра синтез rРНК прекращается, в конце профазы ядрышки исчезают, при конденсации хромосом ядрышковый хроматин в качестве организатора ядрышка входит в SAT-хромосому. После разделения ядра на разрыхляющихся организаторах ядрышка образуются компоненты нового ядрышка. 3.5.4 Ядерная оболочка Ядерная оболочка состоит из двух мембран (каждая толщиной 6–8 нм), между которыми находится перинуклеарное пространство (шириной 10-40 нм). Ядерная оболочка связана с эндоплазматическим ретикулумом и образуется после деления ядра из цистерн ретикулума (используются также обрывки старой ядерной оболочки, разрушенной во время деления). В отличие от других мембран ядерная оболочка обладает видимыми в электронный микроскоп порами (30-100 нм в диаметре), которые занимают около 5% поверхности ядра. Каждая пора с наружной и внутренней стороны окружена кольцевым валиком из 8 сферических рибонуклеопротеидных частиц. В центре поры находится «центральная гранула» – рибонуклеопротеидная частица, которая связана тонкими тяжами с кольцевым валиком и, по-видимому, активно транспортируется в цитоплазму.
Дата добавления: 2014-09-29 | Просмотры: 1530 | Нарушение авторских прав |