|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
ДИССИМИЛЯЦИЯ КАК ИСТОЧНИК ЭНЕРГИИДиссимиляция представляет собой расщепление органических субстратов с использованием их химической энергии.
4.3.1 Обзор процессов диссимиляции При дыханиисубстрат без остаткарасщепляется до бедных энергией неорганических веществ с соответственно высоким выходом энергии. При брожении субстрат разрушается не полностью – до органического конечного продукта, еще богатого энергией, и выход энергии здесь соответственно невелик. Дыхание – аэробныйокислительный процесс, для него необходим кислород. Брожения – анаэробныепроцессы. Высшие животные и растения дышат. Брожение свойственно главным образом микроорганизмам (бактериям, дрожжам), но при недостатке кислорода может встречаться также в клетках высших растений и животных. Важнейшими субстратами для дыхания и большинства брожений служат углеводы.Кроме того, при дыхании могут использоваться жиры и белки. Субстратами для брожения могут также быть спирты, органические кислоты и другие вещества. Из-за малого выхода энергии клетки, осуществляющие брожение, должны расходовать большие количества субстрата, чем дышащие клетки. Дыханиескладывается из двух частичных процессов: 1) постепенного расщепления субстратас отнятием водорода, который связывается с коферментами (символ [Н2]), и 2) постепенного окисления [Н2]в результате переноса его на кислород. Для углеводов:
Молочнокислое брожение(мышцы, бактерии Lactobacillus, Streptococcus): С6Н12О6 ®2СН3СН-ОН-СООН; DG0' = – 200 кДж/моль (4.6) Спиртовое брожение (Saccharomyces и другие дрожжи): С6Н12О6 ®2СН2СН2ОН + 2СО2 ; DG0' = –235 кДж/моль (4.7) Маслянокислое брожение (Clostridium и другие бактерии): С6Н12О6 ® С3Н7СООН + 2СО2 + 2Н2; DG0' = –265 кДж/моль (4.8) У других бактерий встречается муравьинокислое и пропионовокислое брожение.
4.3.2 Пути расщепления углеводов При расщеплении субстрата на него последовательно воздействуют различные ферменты. Некоторые этапы этого процесса при дыхании и брожении идентичны. Расщепление углеводов начинается с гликолиза, после чего пути дыхания и брожения расходятся. Гликолиз – это процесс окислительного расщепления, происходящий в основном веществе цитоплазмы (и клеточного ядра) и ведущий от фруктозо-1,6-бисфосфата к пирувату (пировиноградной кислоте СН3СО-СООН) – промежуточному продукту, имеющему большое значение (рис. 4.4).
Рис. 4.4 – Гликолиз. А. Место в обмене веществ при дыхании и брожении. ФБФ – фруктозо-1,6-бисфосфат; Пир – пируват; Ац-СоА – ацетил-СоА. Б. Химические этапы. В. Окислительная часть цепи гликолиза с фосфорилированием на уровне субстрата. 1 – фруктозо-1,6-бисфосфат; 2 – дигидроксиацетонфосфат; 3 – 3-фосфоглицеринальдегид; 4 – 1,3-дифосфоглицерат; 5 – 3-фосфоглицерат; 6 – 2-фосфоглицерат; 7 – фосфоенолпируват; 8 – пируват (енольная форма и кето-форма)
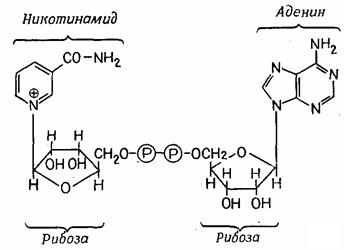
Фруктозо-1,6-бисфосфат сначала расщепляется на два изомерных продукта, находящихся в равновесии между собой. Один из них, 3-фосфоглицеринальдегид, в результате одного этапа окисления превращается в фосфоглицерат, из которого затем образуется пируват. Окисление катализирует дегидрогеназа (фермент, отщепляющий водород). Водород присоединяется к коферменту NAD+ (никотинамидадениндинуклеотид, витамин В3) (рис. 4.5). Восстановленный кофермент NAD-H – один из наиболее универсальных переносчиков водорода. Чтобы энергию окисления можно было сохранить в форме АТР, она используется сначала для «фосфорилирования на уровне субстрата»; при этом образуется высокоэнергетический промежуточный продукт 1,3-дифосфоглицерат. Затем фосфатная группа (вместе с энергией) переносится на ADP, и в результате последний превращается в высокоэнергетическое соединение АТР. Продуктами гликолиза являются пируват, водород в форме NAD-H и энергия в форме АТР.
Рис. 4.5 – NAD+, структурная формула
При разных видах брожения дальнейшая судьба продуктов гликолиза – пирувата и NAD-H – различна. При молочнокислом брожении водород переносится на пируват и в результате сразу образуется лактат (молочная кислота): СН3СО-СООН + NAD-H + Н+ ® СН3СН-ОН-СООН + NAD+ (4.9) При спиртовом брожении пируват сначала декарбоксилируется, т. е. от него отщепляется СО2, а затем промежуточный продукт ацетальдегид восстанавливается в этанол в результате переноса водорода: СН3СО-СООН ® СО2 + СН3СНО (4.10) СН3СНО + NAD-H + H+ ® СН3СН2ОН + NAD+ (4.11) Функция NAD+ и сходных с ним переносчиков водорода состоит в том, чтобы в первой реакции принимать водород (восстанавливаться), а во второй реакции отдавать его (окисляться).
При дыхании водород, получаемый в результате гликолиза, транспортируется к. кислороду. Пируват подвергается окислительному декарбоксилированию при участии мультиферментного комплекса (комплекс белков, осуществляющих несколько ферментативных функций) и нескольких коферментов. Один кофермент принимает отщепленный водород (окисление), а другой – кофермент A (CoA-SH, витамин В5) – присоединяет оставшуюся ацетильную группу (–CO-CH3). CoA-SH содержит реакционно-способные сульфгидрильные группы (–SH) (рис. 4.6).
Рис. 4.6 – Расщепление пирувата – окислительное декарбоксилирование
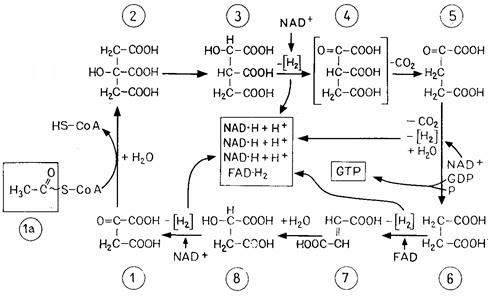
Высокоэнергетическое соединение ацетил-CоА (CoA-S~CO-CH3) («активированная уксусная кислота») является важнейшим промежуточным продуктом клеточного метаболизма, который может иметь различное происхождение и по-разному использоваться. При дыхании ацетильный остаток полностью расщепляется в цикле лимонной кислоты в результате окисления (отнятия Н), декарбоксилирования (отщепления СО2) и гидратации (присоединения Н2О) (рис. 4.7). Общее уравнение: CoA-S~CO-CH3 + 3Н2О ® CoA-SH+ 2CО2+4[H2] (4.12) Перед расщеплением ацетильный остаток связывается с оксалоацета том; в результате образуется цитрат, который постепенно расщепляется, пока снова не остается оксалоацетат; последний вступает в реакцию с новой молекулой ацетил-СоА, и цикл повторяется.
Рис. 4.7 – Цикл лимонной кислоты (цикл трикарбоновых кислот - ЦТК). 1 – оксалоацетат; 1а – ацетил-СоА; 2 – цитрат; 3 – изоцитрат; 4– оксалосукцинат; 5–a-кетоглутарат; 6 – сукцинат (янтарная кислота); 7 – фумарат; 8 – малат (яблочная кислота)
Получаемый в этом цикле водород присоединяют коферменты, прежде всего NAD. Окисление пирувата и цикл лимонной кислоты осуществляются в митохондриях. В основном веществе цитоплазмы возможен еще другой путь расщепления углеводов, без гликолиза и цикла лимонной кислоты, – пентозофосфатный цикл. Глюкозофосфат подвергается окислительному декарбоксилированию до пентозофосфата, а затем 6 молекул пентозофосфата превращаются через ряд промежуточных этапов в 5 молекул глюкозофосфата. Общее уравнение (без фосфата): 6С6Н12О6 + 6Н2О ® 6С5Н10О6 + СО2 + 12 [H2] (4.13) 6С5Н10О5 ® 5С6Н12О6 (4.14) С6Н12О6 + 6Н2О ® 6СО2+ 12[Н2] (4.15)
Дата добавления: 2014-09-29 | Просмотры: 1634 | Нарушение авторских прав |