|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
МИТОХОНДРИИ И ПЛАСТИДЫ
Митохондрии и пластиды представляют собой органеллы эукариотических клеток, сходные по своим функциям, морфологии и, вероятно, происхождению. Они обладают сильно развитой системой внутренних мембран, которая образуется из их оболочки и служит для интенсивного преобразования энергии.
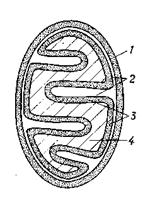
3.7.1 Митохондрии Митохондрии снабжают клетку энергией, которую они накапливают в форме АТР в результате окисления органических веществ (дыхания) – окисления жирных кислот и аминокислот, цикла лимонной кислоты, реакции цепи дыхания и окислительного фосфорилирования. К побочным функциям митохондрий относятся биосинтетические процессы, а также активное накопление ионов. В клетке насчитывается 150–1500 митохондрий, у крупных простейших – до 500 000. Они отсутствуют у ряда паразитических простейших, и в некоторых специализированных клетках (в зрелых эритроцитах). Форма митохондрий – в большинстве случаев от округлой до палочковидной (рис. 3.6), реже нитевидная. Размеры их от 0,5x0,5x1,0 до 1,0x1,0x5,0мкм. Оболочка митохондрий состоит из двух мембран толщиной чаще всего 7–10 нм. Между ними находится перимитохондриальное пространство, а внутри митохондрии – матрикс. Внутренняя мембрана образует многочисленные впячивания; в большинстве случаев это листовидные кристы, у многих простейших и в некоторых клетках млекопитающих – трубочки (тубулы), а у растений – часто кармановидные мешочки.
Рис. 3.6 – Митохондрии. А. Три различных типа внутренней структуры митохондрий: слева – тубулярный, в середине – с кристами, справа – саккулярный. Б. Разделение митохондрий на компартменты: 1 – наружная мембрана; 2 перимитохондриальное (межмембранное) пространство; 3 – внутренняя мембрана; 4 – матрикс
Наружная мембрана (как и другие мембраны эукариотических клеток), в отличие от внутренней мембраны, содержит значительные количества холестерола, но не содержит кардиолипина. Наружная мембрана проницаема для неорганических ионов и для относительно крупных молекул – аминокислот, АТР, сахарозы, промежуточных продуктов дыхания, что можно объяснить наличием туннельных белков с широкими порами. Внутренняя мембрана с кристами по своему составу сходна с бактериальной мембраной: очень богата белком (25% липидов, 75% белков, из них 1/3 периферических и 2/3 интегральных). Она содержит очень мало холестерола; большие количества лецитина и кардиолипина, и имеет другой состав фосфолипидов. Кардиолипин встречается только у прокариот, в митохондриях и в пластидах. Проницаемость внутренней мембраны очень мала, и через нее могут диффундировать только небольшие молекулы. Поэтому в ней имеются транспортные белки для активного транспорта таких веществ, как, например, глюкоза, пируват, метаболиты цикла лимонной кислоты, аминокислоты, АТР и ADP, фосфат, Са2+, и др. В качестве интегральных белков во внутренней мембране и кристах находятся комплексы ферментов, участвующих в транспорте электронов (дыхательная цепь). Периферические мембранные белки – различные дегидрогеназы – окисляют субстраты дыхания, находящиеся в матриксе, и передают отнятый водород в дыхательную цепь. Матрикс содержит промежуточные продукты обмена, некоторые ферменты цикла лимонной кислоты и окисления жирных кислот. Остальные ферменты, участвующие в этих процессах, являются периферическими белками внутренней мембраны. В соответствии со своими функциями митохондрии с высокой интенсивностью биосинтетических процессов богаты матриксом и бедны кристами (например, в печени), в то время как митохондрии, специализированные для выработки энергии (например, «саркосомы» в мышцах), плотно заполнены кристами. Митохондрии содержат в своем матриксе ДНК, РНК (tPHK, rPHK, mPHK, но не 5S- и 5,8S-PHK) и рибосомы (70S у растений и Protozoa, 55S у Metazoa) и способны к репликации ДНК, транскрипции и биосинтезу белка. ДНК, как у прокариот, свободна от гистонов и негистоновых хромосомных белков и представляет собой двухцепочечную кольцевую молекулу. Митохондриальные гены, как и хромосомные, содержат интроны. В каждой митохондрии имеются 2 – 6 идентичных копий молекулы. В митохондриальной ДНК закодированы митохондриальные rРНК и tPHK (с иной первичной структурой, чем у цитоплазматических РНК) и некоторые белки внутренней мембраны. Большинство митохондриальных белков кодируется в хромосомах и синтезируется на цитоплазматических рибосомах. Митохондрии живут только несколько дней, размножаются поперечным делением, но могут также развиваться из промитохондрий. Митохондриальная информация полностью сохраняется и при половом размножении.
3.7.2 Пластиды В зависимости от типа ткани бесцветные пропластиды эмбриональных клеток развиваются в зеленые хлоропласты или в производные от них формы пластид – в желтые или красные хромопласты или в бесцветные лейкопласты. Функция хлоропластов – фотосинтез, т. е. преобразование энергии света в химическую энергию органических веществ, прежде всего углеводов, которые эти пластиды синтезируют из бедных энергией веществ – из СО2 и Н2О. Хлоропласты имеются в клетках, находящихся на свету, у высших растений – в листьях, около поверхности стебля и в молодых плодах. Эти клетки бывают зелеными, если зеленый цвет не маскируется другими пигментами хлоропластов. Пигменты хлоропластов поглощают свет для фотосинтеза. Это в основном хлорофиллы; 70% их составляет хлорофилл а (сине-зеленый), а 30%–хлорофилл b (желто-зеленый) у высших растений и зеленых водорослей и хлорофилл с, d или е у других групп водорослей. Кроме того, все хлоропласты содержат каротиноиды: оранжево-красные каротины (углеводороды) и желтые, реже красные ксантофиллы (окисленные каротины). У красных и синезеленых водорослей встречаются также фикобилипротеиды: голубой фикоцианин и красный фикоэритрин. В клетках высших растений, как и у некоторых водорослей, насчитывается около 10–200 чечевицеобразных хлоропластов величиной всего лишь 3–10 мкм. Оболочка хлоропласта, состоящая из двух мембран, окружает бесцветную строму, которая пронизана множеством плоских замкнутых мембранных карманов (цистерн) – тилакоидов, окрашенных в зеленый цвет (рис. 3.7). Прокариоты не имеют хлоропластов, но у них есть многочисленные тилакоиды, ограниченные плазматической мембраной. В эукариотических растительных клетках тилакоиды образуются из складок внутренней мембраны хлоропласта. Хлоропласты от края до края пронизаны длинными тилакоидами стромы (рис. 3.7), вокруг которых в мелких чечевицеобразных хлоропластах (и только в них!) группируются плотно упакованные, короткие тилакоиды гран (рис. 3.7, А). Стопки таких тилакоидов гран видны в световом микроскопе как зеленые граны величиной 0,3–0,5 мкм.
Рис. 3.7 – Хлоропласты. А. Расположение тилакоидов у высших растений. Б. Хлоропласт в разрезе. В. Модель взаимоотношений между тилакоидами. [По Ohmann (A), Strugger (В)]
Тилакоидные мембраны, которые содержат свыше 40 различных белков, имеют толщину 7–12 нм и очень богаты белком (содержание белка около 50%). Из липидов преобладают гликолипиды. Имеются также фосфолипиды в том числе кардиолипин. В мембранах тилакоидов осуществляется та часть реакций фотосинтеза, с которой связано преобразование энергии, – «световые реакции». В этих процессах участвуют две хлорофиллсодержащие фотосистемы I и II (ФС I и ФС II), связанные цепью транспорта электронов, и продуцирующая АТР мембранная АТРаза. В строме осуществляются биохимические синтезы– темновые реакции фотосинтеза, в результате которых откладываются зерна крахмала (продукт фотосинтеза, пластоглобулы и кристаллы железосодержащего белка). В строме находятся ДНК, mPHK, tPHK, rPHK, 5S-PHK и 70 S -рибосомы. Как и в митохондриях, молекула ДНК замкнута в кольцо, несет гены с интронами и свободна от гистонов и негистоновых хромосомных белков. Имеется от 3 до 30 идентичных копий ДНК на каждый хлоропласт. Молекулы длиннее, чем в митохондриях (40–45, иногда до 160 мкм) и содержат больше информации: ДНК кодирует rРНК и tPHK, ДНК- и РНК-полимеразы, некоторые белки рибосом, а также цитохромы и большинство ферментов темнового процесса фотосинтеза. Однако большая часть белков пластиды кодируется в хромосомах. Лейкопласты – это бесцветные пластиды округлой, яйцевидной или веретенообразной формы в подземных частях растений, семенах, эпидермисе, сердцевине стебля. Они содержат ДНК, зерна крахмала, пластоглобулы, единичные тилакоиды и пластидный центр. Образование тилакоидов и хлорофилла чаще всего либо генетически подавлено (корни, эпидермис), либо тормозится отсутствием света (например, у картофеля: на свету лейкопласты зеленеют и превращаются в хлоропласты). Чаще встречаются амилопласты, образующие крахмал из глюкозы и накапливающие его – главным образом в запасающих органах (клубнях, корневищах, эндосперме и т.п.). Хромопласты являются причиной желтой, оранжевой и красной окраски многих цветков, плодов и некоторых корней. Они бывают округлыми, многогранными, чечевицеобразными, веретеновидными или кристаллоподобными, содержат пластоглобулы (часто в большом количестве), крахмальные зерна и белковые кристаллоиды, не имеют пластидного центра. Тилакоидов в них мало или совсем нет. Пигменты – свыше 50 видов каротиноидов (например, виолаксантин у анютиных глазок, ликопин в помидорах, каротин в моркови) – локализуются в трубчатых или нитевидных белковых структурах или образуют кристаллы. Хромопласты первично нефункциональны. Их вторичная роль состоит в том, что они создают зрительную приманку для животных и тем самым способствуют опылению цветков и распространению плодов и семян. Незрелые пластиды – пропластиды – имеют неправильную форму, окружены двумя мембранами и способны к амебоидному движению. В процессе развития они увеличиваются в размерах, синтезируют крахмальные зерна и кристаллы фитоферритина, и у них образуются трубчатые или листовидные впячивания внутренней мембраны. Для превращения пропластид в хлоропласты необходим свет. Размножение пластид связано с репликацией ДНК и последующим делением пропластиды или хлоропласта надвое.
3.7.3 Филогенез митохондрий и пластид Митохондрии и пластиды занимают в эукариотической клетке совершенно особое положение. Они имеют собственную генетическую систему, размножаются относительно независимо от деления всей клетки и ядра и отграничены от остальной протоплазмы двойной мембраной. Согласно гипотезе эндосимбиоза, они являются потомками прокариот, сходных с бактериями или синезелеными водорослями, которые (вероятно, в результате фагоцитоза) проникли в гетеротрофные анаэробные клетки и стали в них жить, как симбионты. Моделью может служить явление эндоцитоза у некоторых грибов, жгутиковых, амеб, синезеленых водорослей. В пользу этой гипотезы говорят, в частности, следующие общие особенности митохондрий, хлоропластов, бактерий и синезеленых водорослей: 1.Мембрана (внутренняя) содержит кардиолипин; 2.ДНК не связана с гистонами и имеет кольцеобразную форму; 3.Имеются 70S -рибосомы (а не 80S), которые не содержат 5,8S-PHK; 4.Синтез белка начинается с N-формилметионина. РНК хлоропластов и синезеленых водорослей образуют одну группу по родству последовательностей, а РНК митохондрий и бактерий – другую. Обе группы филогенетически далеки от РНК, кодируемой в клеточном ядре. Аргументами против гипотезы эндосимбиоза могли бы служить следующие факты: 1.Большинство белков митохондрий и пластид кодируется в клеточном ядре. 2.Гены митохондрий и пластид содержат интроны, так же как и гены клеточного ядра.
Дата добавления: 2014-09-29 | Просмотры: 1901 | Нарушение авторских прав |