|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
РЕАЛИЗАЦИЯ ГЕНЕТИЧЕСКОЙ ИНФОРМАЦИИ
Генетическая информация закодирована в ДНК. Информация, находящаяся в клеточном ядре, представляет собой генотип. ДНК, содержащаяся в одном наборе хромосом, называется геномом, а внеядерная ДНК (в митохондриях, пластидах и основном веществе цитоплазмы) – плазмоном. У бактерий ДНК в эквиваленте ядра представляет собой геном, а внеядерная ДНК представлена в форме плазмид. Состоящие из ДНК структуры, встречающиеся в основном веществе цитоплазмы у эукариот, так же как и у бактерий, называют плазмидами. Генетическая информация в геноме бактерий и многих вирусов заключена в одной-единственной непрерывной полинуклеотидной цепи. У эукариот генетический материал распределен по хромосомам и в каждой хромосоме тоже образует одну длинную полинуклеотидную цепь. Полинуклеотидные нити ДНК, содержащиеся в хромосомах эукариот, в геноме бактерий и вирусов или плазмидах (у некоторых вирусов – РНК), подразделяются на функциональные отрезки, называемые генами (наследственные задатки). Различают: 1) структурные гены, в которых закодирована информация для синтеза ферментных и структурных белков; 2) гены с информацией для синтеза tPHK; 3) гены с информацией для синтеза rРНК; 4) специфические регуляторные участки, такие как промоторы и операторы; 5) разделяющие участки между генами (спейсеры); 6) участки с неизвестной функцией. Определены последовательности для некоторых полных геномов и для многих частей геномов и сделаны выводы об их функции. Наряду с типичной линейной последовательностью генов в некоторых частях генома существуют отдельные гены, наложенные один на другой так, что небольшие участки нуклеотидной последовательности принадлежат различным генам. Гены отделены друг от друга спейсерами. Кроме того, имеются участки, «распознаваемые» определенными молекулами, такие как промотор и оператор, для регуляции активности генов. У эукариот помимо структурных генов, которые расположены в хромосомах линейно, существуют участки с повторяющимися последовательностями.
5.1 ДЕЙСТВИЕ ГЕНОВ Первым шагом на пути к формированию признака является транскрипция – «переписывание» нуклеотидных последовательностей определенных участков ДНК в форме mPHK. После этого РНК-транскрипт (пре-РНК) может модифицироваться в результате посттранскрипционных процессов. На рибосомах осуществляется трансляция – перевод нуклеотидной последовательности mPHK в аминокислотную последовательность полипептида. Наряду с многими белками для этого процесса необходимы также tPHK и rРНК. Те или иные признаки клетки или организма появляются благодаря тому, что образуются специфические структурные белки или же ферменты, ответственные за определенные этапы процессов синтеза или распада. В результате из предшественников через промежуточные этапы образуются конечные продукты, от которых и зависит проявление специфических признаков, т. е. функциональных способностей (рис. 5.1).

Рис. 5.1 – Реализация генетической информации (схема)
5.1.1 Транскрипция При транскрипции рибонуклеозиды – цитидин, гуанозин, уридин и аденозин – С, G, U и А, синтезированные в процессе клеточного метаболизма в форме рибонуклеозидтрифосфатов (rNTP) СТР, GTР, UТР и АТР, пристраиваются к комплементарным основаниям ДНК, а именно C к G, G к C, U к А и А к Т. Транскрипция идет от начала транскрипционной единицы до ее конца. ДНК-матрицей служит так называемая кодогенная цепь ДНК, в которой транскрипционные единицы транскрибируются в направлении 3'→5'. Рибонуклеозидтрифосфаты (rNTP) с помощью ферментов – ДНК-зависимых РНК-полимераз – связываются (с последующим отщеплением пирофосфата) в направлении 5' → 3' в цепь РНК (рис. 5.2, А). Этот фермент должен не только обеспечивать правильное образование пар и связывание rNTP в цепь, но и находить надлежащее место начала транскрипции, выбирать кодогенную цепь и разделять две цепи ДНК около тех пар оснований, где происходит транскрипция. Синтез РНК заканчивается на терминаторном участке ДНК. Транскрипция активного гена осуществляется так, что на транскрипционной единице одновременно находится много молекул полимеразы с растущими цепями РНК (рис. 5.2, Б). У эукариот для транскрипции ядерной ДНК существуют три различные РНК-полимеразы; структурные гены транскрибируются полимеразой II.
Рис. 5.2 – Транскрипция. А. Синтез mPHK в начальной части гена. Б. Повторный старт синтеза mPHK на гене. В. Транскрибируемый участок генетического материала
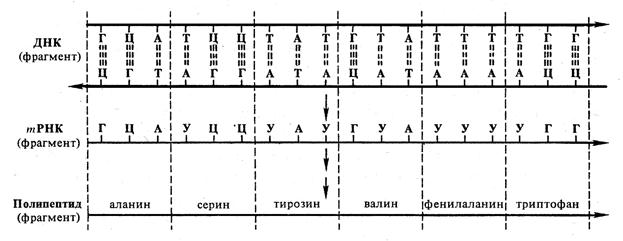
В одной определенной фазе жизненного цикла транскрипции подвергается лишь около 10% структурных генов; остальные гены неактивны, но могут стать активными в других фазах. Какие гены активны, зависит от жизненного цикла клетки, от ее дифференцировки, от стадии онтогенетического развития организма и от факторов внешней среды. У эукариот транскрипция происходит в клеточном ядре, а трансляция – в цитоплазме на рибосомах. Пре-mPHK в форме рибонуклеопротеидных частиц попадает в цитоплазму и претерпевает ряд изменений, которые называют посттранскрипционными процессами. Как у прокариот, так и у эукариот первичный транскрипт (пре-mPHK) в большинстве случаев длиннее, чем последовательность нуклеотидов, соответствующая конечному продукту (полипептиду, tPHK, гРНК). Готовая mPHK начинается с вводной последовательности (лидера), затем следуют участок, несущий информацию для генного продукта, и концевая последовательность (трейлер). У эукариот часто встречаются многообразные различия между первичным транскриптом и РНК, поступающей для трансляции. После транскрипции могут происходить следующие изменения (в совокупности называемые также процессингом: а) образование «колпачка» (метилирование и формирование специальной структуры на 5'-конце); б) метилирование внутренних оснований; в) разделение всей последовательности на части или отщепление определенных частей; г) удаление внутренних участков и соединение оставшихся последовательностей – сплайсинг; д) добавление poly (А) к 3'-концу. Образование колпачка и внутреннее метилирование пре-mРНК типично для эукариот. У эукариот в последовательности нуклеотидов ДНК с информацией для последовательности аминокислот некоторых полипептидов включены отрезки, не содержащие информации, так называемые интроны. Сначала образуется транскрипт всей последовательности (пре-mPHK), а затем происходит сплайсинг и информативные отрезки объединяются в одну непрерывную последовательность – mРНК. Предполагают, что часть посттранскрипционных процессов имеет значение для регулирования действия генов. Для начала транскрипции большое значение имеет так называемый промотор. Это предшествующая гену последовательность состоит примерно из 80 нуклеотидов, которую узнает и с которой связывается фермент РНК-полимераза. 5.1.2 Трансляция При трансляции нуклеотидная последовательность tРНК переводится в аминокислотную последовательность полипептидной цепи. Ключом для перевода последовательности нуклеотидов в последовательность аминокислот служит генетический код. Как при переводе с одного языка на другой необходимо знать слова, так и здесь следует знать, как каждая аминокислота закодирована в нуклеотидах. В tРНК имеется 4 различных нуклеотида: С, G, U и А. В биосинтезе белка участвуют 20 аминокислот. Чтобы их закодировать, необходимо по меньшей мере 20 различных знаков. При четырех различных нуклеотидах это возможно в случае составления знаков из трех нуклеотидов; таких знаков может быть 43 = 64. Три нуклеотида, образующие кодовый знак, называют триплетом. Генетический код удалось выяснить благодаря работам исследовательских групп Ниренберга и Очоа (табл. 5.1, рис. 5.3). Результаты этих работ явились одним из самых значительных шагов в понимании жизненных процессов. Эти результаты можно кратко резюмировать следующим образом: 1. Генетический код представляет собой триплетный код. Триплет mРНК получил название кодона. 2. Генетический код является вырожденным кодом, т. е. одной аминокислоте, как правило, соответствует более чем один кодон. В кодонах для одной аминокислоты первые два нуклеотида чаще всего одинаковы, а третий варьирует. 3. Нуклеотидная последовательность считывается в одном направлении подряд, триплет за триплетом. Кодоны не перекрываются. 4. AUG представляет собой стартовый кодон. 5. UAG (amber), UAA (ochre) и UGA (opal) – кодоны-терминаторы. 6. Генетически код универсален, он един для всех организмов и вирусов. Таблица 5.1 – Генетический код (кодоны tРНК для аминокислот)
Рис. 5.3 – Генетический код. Соотношение последовательности триплетов ДНК, РНК и мономеров (аминокислот) в белковой молекуле
Трансляция осуществляется на рибосоме. tРНК прикрепляется к малой субчастице, связывание аминокислот происходит на большой субчастице. Кроме информации для аминокислотной последовательности, tРНК содержит начальную (лидер) и концевую (трейлер) последовательности. Часть начальной последовательности необходима для связывания с малой субчастицей рибосомы. «Колпачку» у эукариот приписывают роль в узнавании рибосомы и в связывании с малой субчастицей. Трансляция начинается со стартового кодона AUG (рис. 5.4). На малой субчастице есть особый участок (позиция 1), где триплет подготавливается к трансляции. Так как генетический код не имеет «запятых», начало считыва-
А
Б
Рис. 5.4 – Трансляция. А. Обединение субчастиц рибосомы на mРНК (схема). Б. Образование белков на рибосоме
ния должно определяться точно, и для этой цели служит стартовый кодон AUG. Затем на то же место для считывания непрерывно друг за другом встают последующие триплеты. Роль посредника между кодоном mРНК и аминокислотой играет tPHK. Для каждой аминокислоты существует, по меньшей мере, одна tPHK. Она выполняет в основном две задачи: выбирает из 20 аминокислот специфичную для нее и находит с помощью своего антикодона (рис. 2.8. и 5.4) соответствующий кодон mРНК по принципу спаривания оснований. Та tPHK, которая, подойдя к малой субчастице, образует связь кодон-антикодон, одновременно передает свою аминокислоту в аминоацильный участок большой субчастицы. К кодону AUG «подходит» антикодон только той tPHK, которая переносит метионин (Met-tPHK). Поэтому прежде всего к рибосоме доставляется метионин. Затем одновременно кодон AUG переходит из позиции 1 в позицию 2 на малой субчастице, а Met-tPHK – на пептидильный участок большой субчастицы. На малой субчастице в позиции 1 стоит теперь следующий кодон (на рис. 5.4 – UUU), готовый к связыванию с антикодоном. К кодону UUU «подходит» антикодон Phe-tPHK, и в аминоацильный участок большой субчастицы попадает аминокислота фенилаланин. Карбоксильная группа метионина, которая раньше была связана с tPHK, присоединяется к аминогруппе фенилаланина, и образуется дипептид. Met-tPHK освобождается и готова теперь связать новую молекулу метионина. Когда второй кодон, UUU, переходит в позицию 2, позиция 1 освобождается для следующего кодона (на рис. 5.4 – ССС). Снова происходит спаривание оснований кодона и антикодона. На аминоацильный участок большой субчастицы попадает аминокислота пролин. Образуется пептидная связь между фенилаланином и пролином. Таким образом считывается триплет за триплетом. Последовательность кодонов в tРНК определяет последовательность аминокислот в полипептиде. Образование пептидных связей прекращается, когда в участке трансляции появляется кодон-терминатор tРНК. Для него не существует tPHK, и в аминоацильный участок не попадает никакая аминокислота. Обе субчастицы объединяются в способную функционировать рибосому только тогда, когда к малой субчастице уже прикрепилась mРНК, а к последней присоединилась Met-tPHK. Эту начальную фазу трансляции называют инициацией. Связывание последующих аминокислот в полипептид происходит в фазе элонгации (удлинения цепи). В фазе терминации синтез полипептидной цепи заканчивается при участии освобождающих цепь факторов. tРНК одновременно считывается несколькими рибосомами («полисомы»). Еще до того как она будет полностью транслирована, ее начало попадает на следующую рибосому и там тоже транслируется. На рибосоме правильное размещение аминокислот в полипептидной цепи осуществляется благодаря спариванию оснований между антикодоном tPHK и кодоном tРНК. За связывание аминокислоты с соответствующей tPHK ответствен специфический фермент – синтетаза. Каждая аминокислота имеет свою специфическую синтетазу (или несколько таких синтетаз). Все tPHK имеют акцепторный конец, к которому присоединяется аминокислота (см. рис. 2.8). Присоединение аминокислот к специфическим tPHK – необходимая предпосылка безошибочного хода биосинтеза белков, так как каждая аминокислота, связанная с tPHK, включается в пептидную цепь в соответствии с антикодоном этой tPHK.
Дата добавления: 2014-09-29 | Просмотры: 2090 | Нарушение авторских прав |