|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
АССИМИЛЯЦИЯ
Ассимиляция – это превращение чужеродных веществ в компоненты собственного организма. Автотрофная ассимиляция бывает двух видов – фотосинтез и хемосинтез (табл. 4.4). Зеленые растения, синезеленые водоросли и некоторые бактерии, синтезирующие органические вещества из неорганических – «первичные продуценты», имеют огромное значение для жизнедеятельности всех живых существ. Гетеротрофная ассимиляция остальных организмов – сравнительно более простой процесс превращения одних органических веществ в другие.
Таблица 4.3 – Последовательности реакций в процессе дыхания и их энергетический выход
* – в скобках показано количество АТР, образуемое при дыхательном метаболизме
Таблица 4.4 – Формы автотрофной ассимиляции
Так как органические вещества представляют собой соединения углерода, то решающее значение имеет ассимиляция углерода. Это процесс восстановления, который ведет от максимально окисленного исходного вещества СО2 к менее окисленным продуктам, таким как углеводы [(СН2О)n]. У зеленых растений и сине-зеленых водорослей источником необходимых для восстановления электронов служит вода (Н2О), которая при отнятии электронов окисляется. Автотрофные бактерии не способны к окислению воды, им нужны другие доноры электронов – S0, H2S, NH3, NO2–, Fe2+. Потребность в энергии удовлетворяет свет (фотосинтез, или окисление неорганических соединений – хемосинтез).
4.4.1 Фотосинтез (общий обзор) Фотосинтез – это преобразование энергии света в химическую энергию. Такое преобразование происходит в тилакоидах. Химическая энергия накапливается прежде всего в форме АТР и [Н2] (водород, связанный с коферментом). Для облигатных автотрофов (зеленые бактерии, пурпурные серобактерии, многие сине-зеленые водоросли) фотосинтез – единственный источник энергии: у них нет процессов диссимиляции, поставляющих АТР. В зеленых клетках высших растений в цитоплазму переходят большие количества АТР и [Н2]. Значительная часть последнего (в форме NAD-H+H+) попадает в митохондрии и там окисляется в цепи дыхания для дополнительного синтеза АТР. У высших растений большая часть АТР и [Н2] используется для синтеза углеводов из СО2. Таким образом, фотосинтез включает преобразование энергии (световой процесс) в тилакоидах хлоропластов и превращения веществ (ассимиляция углерода – темновой процесс) в строме хлоропластов:
Восстановитель [Н2] (е–+Н+) образуется при расщеплении воды за счет энергии света ( фотолиз ), при котором выделяется О2. АТР синтезируется при прохождении электронов по цепи транспорта электронов. Переносчиком водорода служит NADP (никотинамидадениндинуклеотидфосфат), который по сравнению с NAD содержит на один фосфатный остаток больше. NADP-H + + H+ и АТР направляются в темновой процесс, где водород и энергия используются для синтеза углеводов из СО2, а затем NADP+ и ADP снова используются в световом процессе. Другие органические вещества (жирные кислоты или аминокислоты) могут быть побочными продуктами фотосинтеза или же вторично образуются из углеводов. На каждые 6 молей поглощенного СО2 выделяется 6 молей О2. Коэффициент ассимиляции AQ – отношение О2/СО2– при биосинтезе углеводов равен 1. Для восстановления одной молекулы СО2 необходимо около 9 квантов света, так что на 1 моль СО2 должно приходиться 9 молей квантов. Так как 1 моль квантов красного светасодержит 172 кДж, затрата энергии равна около 9×172 кДж на 1 моль СО2, т. е. 6×9×172 кДж=9288 кДж на 1 моль С6Н12О6. Согласно приведенному выше уравнению, из этого количества 2875 кДж связывается в форме химической энергии (в углеводе); остаток (около 70%) – потери.
4.4.2 Преобразование энергии в фотосинтезе (световой процесс) В расчете на 1 молекулу О2 (или 1 молекулу СО2) световой процесс можно представить так: 2Н2О + Световая энергия ® О2 + 2[Н2] + Энергия АТР, или подробнее: Свет 2Н2О+ 2NADP+ + n(ADP + фосфат) ® О2 + 2(NADP-H + Н+) + nАТР (4.23)
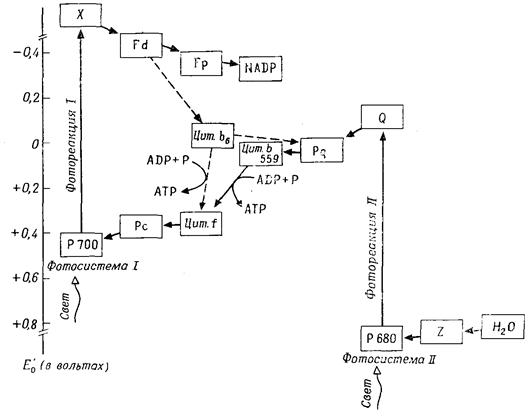
Таким образом, световой процесс представляет собой перенос водорода (электронов и протонов) с одной окислительно-восстановительной системы (Н2О/0,5·О2+2Н++2е–; Е¢0=+0,81В) на другую (NADP-H+/NADP++2H++2e–; E0= = – 0,32В). Однако перенос электронов от положительного потенциала к отрицательному– процесс эндергонический, он требует затраты энергии. Только для этого и нужна при фотосинтезе энергия света. Таким образом, первичное фотохимическое событие – это перенос электронов против градиента окислительно-восстановительного потенциала(ОВП) за счет энергии света. Для этого переноса используется цепь транспорта электронов. На большинстве этапов электроны перемещаются здесь «вниз»по градиенту ОВП без затраты энергии и без света. И только два этапа осуществляются противградиента ОВП за счет световой энергии; будучи фотохимическими реакциями, эти этапы не зависят от температуры и протекают даже при минимальных температурах. Фотохимическое действие могут оказывать только те кванты света, которые поглощаются пигментами. Тилакоиды содержат следующие пигменты, связанные с белками: хлорофиллы, каротиноиды (каротины и ксантофиллы), а у красных и синезеленых водорослей также фикобилипротеиды. Свет поглощают все пигменты, но только фотосинтетически активные пигменты (хлорофилл а у растений и синезеленых водорослей, бактериохлорофилл у бактерий) выполняют при этом фотохимическую работу (транспорт электронов). Добавочные пигменты (хлорофилл b, каротиноиды, фикобилипротеиды) передают поглощенную энергию активным пигментам без существенных потерь. Хлорофиллы поглощают свет в синей и красной областях спектра, каротиноиды – в синей и сине-зеленой областях. В зеленой и желтой областях светне поглощается (исключение составляют красные и сине-зеленые водоросли) и фотосинтеза не происходит. При поглощении светового кванта молекула пигмента возбуждается, т.е. на короткое время переходит в высокоэнергетическое, возбужденное состояние. При ее возвращении в исходное состояние выделяется энергия, за счет которой может совершаться различная работа. Хлорофилл может иметь различные возбужденные состояния. При возвращении в исходное состояние энергия может выделяться в виде флуоресценцииили тепла, передаваться в качестве возбуждающей энергиидругим молекулам или использоваться для фотохимической работы. В тилакоидных мембранах молекулы пигментов расположены вместе с белками и другими компонентами в двух различных комплексах – фотосистеме I и фотосистеме II (ФС I и ФС II) (рис. 4.11). Каждая фотосистема содержит, во-первых, 1 молекулу «пигмента реакционного центра»(ПРЦ,
Рис. 4.11 – Фотоперенос электронов. Линейная и циклическая цепи транспорта электронов
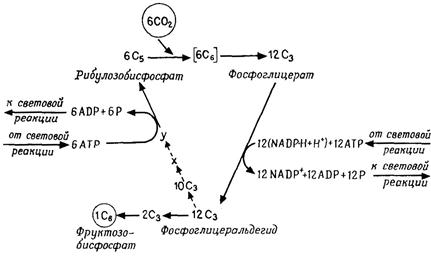
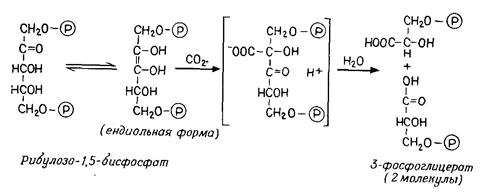
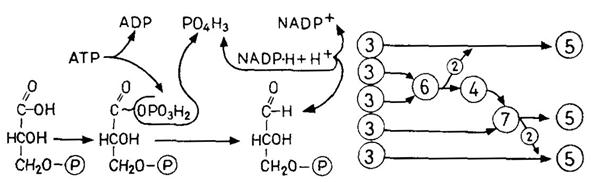
хлорофилл а), которая после поглощения света (возбуждения) выполняет фотохимическую работу (перенос электронов), и, во-вторых, множество молекул «пигментов-антенн»или «коллекторов»(хлорофиллы а и b,каротиноиды), передающих поглощенную энергию ПРЦ и возбуждающих его. Каждая частица ФС I и ФС II содержит 200–400 молекул хлорофилла. ФС I имеет в качестве ПРЦ пигмент 700 (хлорофилл а1): две молекулы хлорофилла, которые благодаря взаимодействию диполь-диполь возбуждаются легче, чем хлорофилл-мономер. ПРЦ в ФС II представляет собой пигмент 680 (хлорофилл а2). ФС II содержит особенно много хлорофилла b. Фотохимическая работа пигмента реакционного центра осуществляется следующим образом: возбужденная молекула пигмента (Хл) отдает валентный электрон акцептору электронов с отрицательным окислительно-восстановительным потенциалом (ОВП); образующийся при этом пигментный катион (Хл+) отнимает электрон от донора электронов с положительным ОВП. Таким образом, электроны переходят с более низкого энергетического уровня на более высокий против градиента ОВП: ФС I переводит электроны с Е'0 + 0,4 В на Е'0 – 0,4 В (фотореакция I), а ФС II – с Е'0+0,8 В на Е'0 –0,15 В (фотореакция II). Цепь транспорта электронов идет от Н2О (Е'0=+0,81 В) через обе фотосистемы к NADP (Е'0= – 0,32 В). В фотореакции II (в ФС II) и фотореакции I (в ФС I) электроны последовательно два раза поднимаются «в гору», каждый раз за счет энергии одного кванта света (эндергонические процессы); на промежуточном этапе они спускаются «под гору» (экзергонический процесс), при этом образуется АТР. Этот путь называется линейным (нециклическим) фотопереносом электронов или Z-фотосинтезом. Донор электронов Н2О отдает электроны переносчику электронов Z (Mn-протеиду), от которого они через пигмент 680 переходят к акцептору электронов в ФС II – «гасителю» Q неизвестной химической природы (фотореакция II). Следующий переносчик электронов пластохинон (Pq) в химическом и функциональном отношении сходен с убихиноном и, так же как и последний, растворен в липидной фазе мембраны. Далее идет цитохром b559 – железопорфирин, как и все цитохромы; он является еще компонентом частиц ФС II, тогда как пластоцианин (Рс – Сu-протеид, переносящий электроны) находится в электронотранспортных частицах тилакоидной мембраны. От Рс электроны через пигмент 700 передаются акцептору электронов в ФС I – веществу X (фотореакция I) – и далее ферредоксину (Fd – белку, содержащему железо и серу) и приобретают весьма высокую энергию, так как Fd обладает чрезвычайно низким окислительно-восстановительным потенциалом (Е'0 = –0,43 В). Флавопротеид (Fp) с FAD в качестве ко-фермента осуществляет затем перенос электронов на NADP. Фотолиз воды. При линейном фотопереносе электронов используются кванты света и Н2О. В результате отрыва электронов под действием света (фотоокисление) соответствующие молекулы воды распадаются (фотолиз воды), образуя протоны и О2. Этот кислород, освобождающийся при фотосинтезе, происходит из Н2О, а не из СО2. Линейный фотоперенос электронов поставляет два продукта: АТР и NADP-H + H+. Освобождение протонов при фотолизе Н2О уравновешивается использованием их при образовании NADP-H + H+. Циклический фотоперенос электронов. Фотосистема I может быть закорочена. Тогда возникает круговой ток электронов при участии цитохрома b6, который вместе с цитохромом f (и Рс) присутствует в электронотранспортных частицах тилакоидных мембран. Электроны перекачиваются «в гору» с помощью одиночных квантов света, а при их обратном «спуске» образуется АТР. Циклический фотоперенос электронов не включает фотолиза Н2О и не служит источником NADP-H. Здесь только один продукт – АТР. Синтез АТР в данном случае называют циклическим фотофосфорилированием в отличие от нециклического при линейном фотопереносе электронов. Циклическая цепь имеется во всех участках тилакоидов – и в гранах, и между гранами, а фотосистема II и, следовательно, линейная цепь, – вероятно, только в гранах. 4.4.3 Превращения веществ при фотосинтезе (темновой процесс) В темновом процессе при использовании продуктов светового процесса (NADP-H и АТР) из СО2 синтезируется углевод (рис. 4.12). В расчете на 1 молекулу СО2 (или О2) СО2 + 2[Н2] + Энергия АТР ® (СН2О) + Н2О, или подробнее: СО2 + 2(NADP-Н + Н+) + 3АТР ® (СН2О) + 2NADP+ + Н2О +3(ADP+фосфат), (4.24) где (СН2О) означает 1/6 молекулы глюкозы. Главный путь ассимиляции СО2 – цикл Кальвина – циклический процесс, в который вводится СO2 и из которого выходит углевод (рис. 4.12, А). Процесс можно разделить на три фазы: Фаза карбоксилирования (рис. 4.12, Б). СО2, связываясь с рибулозобисфосфатом (фосфатом сахара с пятью атомами С), образует две молекулы фосфоглицерата (по 3 атома С – С3). Эту реакцию катализирует рибуло-зобисфосфаткарбоксилаза. Фаза восстановления. Фосфоглицерат при участии NADP-H (восстановитель) и АТР (донор энергии) восстанавливается до 3-фосфоглицеральдегида. Эта последовательность реакций (рис. 4.12, В) представляет собой обращение окислительных этапов гликолиза. Фаза регенерации. Каждая шестая молекула фосфоглицер-альдегида выходит из цикла (рис. 4.12, Г), и из этого вещества образуется фруктозо-1,6-бисфосфат (С6); из последнего в свою очередь синтезируются глюкоза, сахароза, крахмал и т. д. Из остальных молекул фосфоглицеральдегида (С3) при участии новых молекул АТР регенерируется рибулозобисфосфат (5С3®ЗС5); в качестве промежуточных продуктов образуются различные фосфаты сахаров например, С4 – эритрозо-, С6 – фруктозо-, С7 – седогептулозофосфат. С окончанием этой фазы цикл замыкается.
Рис. 4.12 – Цикл Кальвина. А. Суммарная схема. Вверху – фаза карбоксилирования; справа – фаза восстановления; слева – фаза регенерации. Б. Фаза карбоксилирования; нестабильный промежуточный продукт (С6) подвергается гидролизу. В. Фаза восстановления (с 1,3-дифосфоглицератом в качестве промежуточного продукта, ср. рис. 4.4, В). Г. Фаза регенерации. Большие кружки – промежуточные продукты; маленькие кружки – переносимые фрагменты молекул; цифрами в кружках указано число атомов. (А, Б и Г по Libbert.) Ферменты цикла находятся в строме хлоропласта, а рибу-лозобисфосфаткарбоксилаза – также на наружной стороне тилакоидных мембран.
Дата добавления: 2014-09-29 | Просмотры: 2526 | Нарушение авторских прав |