|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Причины наступления родов и регулярные механизмы сократительной деятельности матки
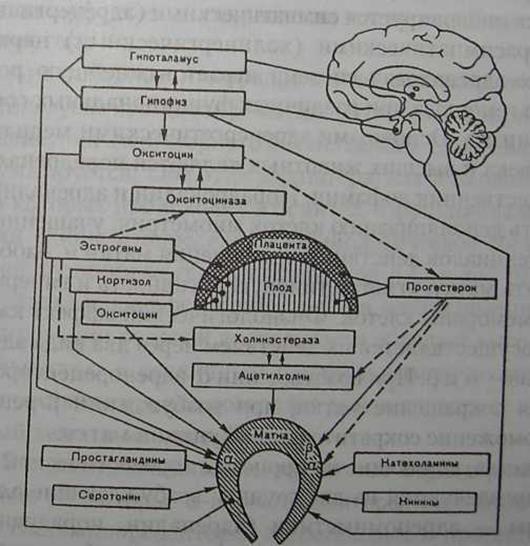
Родовой акт - это сложный многозвеньевой физиологический процесс, который возникает и завершается в результате взаимодействия многих систем организма Регуляция моторной функции матки в организме беременной осуществляется нервными и гуморальными путями (рис. 14). При этом центральной нервной системе принадлежит решающая роль в создании оптимальных условий для развития и нормального течения беременности и родов В ос нове возникновения и развития родовой деятельности лежит безусловный цепной рефлекс. Рефлексы начинаются с рецеп-
Рис. 14. Схема регуляции сократительной деятельности матки. Сплошные стрелки — активация (стимуляция), пунктирные (подавление)
торов матки, которые воспринимают раздражение от плодного яйца и плода и играют большую роль во взаимосвязи функции материнского организма в соответствии с состоянием плода. При изменении информации, идущей от плода, наблюдаются рефлекторные изменения тонуса и моторики матки. Начало родов следует рассматривать как результат постепенной интеграции связи морфологических, гормональных, биохимических и биофизических состояний. В процессе родов имеет значение раздражение рецепторов матки и родовых путей. По мере вовлечения в процесс новых рецепторов меняются сила и частота сокращений матки, а в дальнейшем присоединяются сокращения поперечно- полосатой мускулатуры (потуги). Характер и степень выраженности различных рефлекторных реакций во многом зависят от воздействия на нервную систему гуморальных и гормональных факторов, а также от тонуса симпатического и парасимпатического отделов вегетативной нервной системы Матка иннервируется симпатическими (адренергическими) и парасимпатическими (холинергическими) нервами. Симпатико-адреналовая система играет важнейшую роль в регуляции гемостаза при различных функциональных состояниях организма. Основными адренергетическими медиаторами у человека и высших животных являются норадреналин и его предшественник дофамин. Норадреналин и адреналин могут вызвать деполяризацию клеток миометрия, учащение разрядов потенциалов действия и сокращения матки и, наоборот, расслаблять матку, угнетая типовую активность и гиперполяризацию мембраны клеток. Физиологический эффект катехо-ламинов осуществляется их действием через два вида адрено-рецепторов - а и b. При возбуждении а-адренорецепторов наблюдается сокращение матки, при возбуждении р-рецепторов - торможение сократительной функции матки. Средства, влияющие на процессы адренергической передачи, подразделяются на две группы: возбуждающие адрено-рецепторы - адреномиметики (адреналин, норадреналин, эфедрин, ритодрин, лизатон, алупент и др.) и блокирующие адренорецепторы - адренолитики или адреноблокаторы (фетоламин, пропранолол, индерал, обзидан и др.). При физиологическом течении беременности преобладает тонус адренергической системы. В процессе родов наблюдаются резкие колебания тонуса того или иного отдела вегетативной нервной системы. Во время сокращений матка испытывает резко выраженное влияние со стороны холинергичес-кой системы, а во всех остальных частях организма проявляется отчетливо выраженная симпатикотония (повышение артериального давления, тахикардия, расширение зрачков, усиление потоотделения, усиление дермографизма и др.). При физиологически протекающих родах под воздействием определенного количества медиаторного ацетилхолина наступают сокращения маточной мускулатуры, разрушение ацетилхолина холинэстеразой сопровождается постепенным падением волны сокращения. Следующие сокращения матки возникают при появлении новой порции ацетилхолина. Если нарушается механизм своевременного, ритмического разрушения ацетилхолина, то вследствие задержки последнего в соприкосновении с ганглиозными клетками очень скоро эффект возбуждения нервно-мышечного аппарата сменяется депрессивным эффектом — сокращения матки ослабевают или прекращаются. Ацетилхолин и норадреналин, оказывая тонотропное влияние, усиливают тонус матки, создают благоприятный фон для действия медиаторного ацетилхолина. Действие их на матку суммируется. Оба этих медиатора, как и нервные структуры, в которых происходит их синтез, в функциональном отношении едины и определяют физиологическое состояние моторной функции матки (Персианинов Л.С. и др., 1975). Матка как эффекторное звено в возникновении и функционировании родовой деятельности обладает высокой чувствительностью к различным гуморальным веществам, гормонам и растяжению, что позволяет рассматривать ее как своеобразный хемо- и механорецептор. В миометрии выявлены различные медиаторные и гормональные рецепторы: а-адренорецепторы, серотонин, м-холино- и гистаминорецепторы, эстрогеновые и прогестероновые. окситоциновые, простагландиновые рецепторы. Важно подчеркнуть, что в шейке матки имеются рецепторы к простагландину Е2, тогда как в теле матки — к простагландину F2a, Чувствительность рецепторов матки зависит от гормонального фона, в основном от соотношения половых стероидных гормонов — эстрогенов и прогестерона. M.L.Casey и Р.С. Mac Donald (1988) считают целесообразным различать термин утеротропины и утеротонины. Под утеротропинами они понимают вещество или вещества (агенты), которые готовят матку к родам (размягчение и созревание шейки, появление связей между клетками миометрия, увеличение количества окситоциновых рецепторов в миометрии, возрастание контрактильной чувствительности миометрия к утеротонинам). Утеротонин — термин который используется для веществ, как например постагландины или окситоцин, которые вызывают сокращения миометрия. Возможно, что утеротонин, также является и утеротропином. Среди утеротропинов особое место занимают эстрогенные гормоны и прогестерон. Эстрогенные гормоны обеспечивают рост мышечной и соединительной ткани миометрия, усиливают синтез актомио-зина, способствуют накоплению гликогена и фосфорных соединений, снижают потенциал покоя, увеличивают накопление ионов кальция, стимулируют а-адренорецепторы, повышают чувствительность матки к окситотическим веществам, поддерживают интенсивный кровоток в матке, повышают синтез простагландинов (Михайленко Е.Т., 1978). Эстрогены играют несомненную роль в повышении возбудимости матки, способствуют созреванию шейки и подготовке мягких родовых путей к родам. Они подавляют активность окситоциназы, предохраняя тем самым эндогенный окситоцин от разрушения. Увеличение окситотической активности сопровождается снижением активности холинэстеразы и увеличением концентрации свободного ацетилхолина, оказывающего мощное кон-трактильное действие. Повышая чувствительность к окситотическим веществам миометрия и обеспечивая его пластическим материалом, эстрогены способствуют физиологическому течению родового акта. Прогестерон повышает мембранный потенциал, блокирует транспорт ионов натрия и стабилизирует клеточную мембрану, влияя на тонус матки, амплитуду и частоту схваток. Мембранный потенциал клеток миометрия и концентрация прогестерона в области прикрепления плаценты выше, чем в клетках других отделов матки (Csapo A., 1969). Выявлена зависимость между локализацией плаценты и длительностью родов. Так, при расположении плаценты в области дна матки сокращения ее слабее и слабость родовой деятельности отмечается чаще, чем при локализации плаценты в теле и нижнем сегменте (Давыдов С.Н. и соавт., 1976). Полагают, что прогестерон оказывает действие на гипоталамо-гипофизарную систему, подавляя экскрецию окситоцина. Большую роль в развитии родовой деятельности играют соотношения между эстрогенами и прогестероном. Чем ниже прогестерон — эстрогеновый индекс, тем выше готовность организма к родам (Чернуха Е.А., 1982). Определенное значение в развитии родовой деятельности придают кортикостероидам. Причина повышенной концентрации кортикостероидов связана с увеличением их синтеза надпочечниками матери и плода, а также повышенной их продукцией в плаценте. Полагают, что плод является источником сигнала для начала родов, который заключается во внезапном подъеме уровня кортизола в периферической крови плода. Подъем уровня кортизола вызывает увеличение содержания эстрогенов и простагландинов. Выявлено повышение уровня АКТГ и кортизола в крови пуповины в родах. Однако неясно, является это результатом родового стресса или стимуляция функции гипофизарно-надпочечниковой системы плода предшествует наступлению родов (Баграмян Э.Р.и Цибульская И.С, 1976; Захарова О.И., 1986; Poulakka J. et al., 1982). В регуляции моторной функции матки наряду с гормональным влиянием определенная роль принадлежит серотонину, кининам, ферментам - веществам-посредникам, через систему которых гормоны воздействуют на биоэлектрический и пластический процесс в миометрии. Серотонин — биологически активный амин, один из медиаторов нервного возбуждения, способствующий увеличению содержания кальция в мышце матки. Действие серотонина на матку может осуществляться Двумя путями — непосредственно на миометрии через а-рецепторы и через центральную нервную систему путем усиления выработки окситоцина нейрогипофизом. Концентрация серотонина находится в прямой зависимости от активности мионоаминоксидазы (МАО), разрушающей его. По мере увеличения срока беременности активность МАО постепенно снижается, а содержание серотонина в крови, миометрии и плаценте в связи с этим увеличивается и достигает максимума к моменту родов, что свидетельствует о его непосредственном участии в родовом акте. Подтверждением этого является тот факт, что серотонин с успехом используют для возбуждения и стимуляции родовой деятельности (Бакшеев Н.С. и Орлов Р.С., 1976; Михайленко Е.Т. и соавт., 1980; Urban, 1975). Морфологические исследования миометрия показали, что его поверхностный слой активного участия в родовой деятельности не принимает. В глубоких слоях миометрия выявлена высокая активность веществ, увеличено содержание РНК, белков, гликогена, что свидетельствует о повышении окислительно-восстановительных процессов. При нормальной родовой деятельности в миометрии происходят усиление синтеза белков, накопление РНК, снижение уровня гликогена и увеличение активности энзимов пентозного ряда. При слабости родовых сил биоэнергетические процессы нарушаются, о чем свидетельствует снижение содержания РНК, основного белка, SH-групп, липидов и углеводсодержащих соединений, снижается активность ферментов (Персианинов Л.С. и соавт., 1975). Начало родов при доношенной беременности объясняют ускоренным синхронным образованием утеротропин-утеротонинов и изменениями в матке, которые и ведут к началу родов. Имеется множество натуральных соединений, которые вызывают сокращения матки. К утеротонинам относятся простагландины, оксшпоцин, ангиотензин II, аргивин, вазопрессин, брадикинин. Некоторые из них образуются в матке во время беременности в децидуальной ткани, оболочках плодного яйца, как например ПГР2а, ПГЕ2 и аккумулируются в амниотической жидкости. Простагландины могут воздействовать через кровоток матери и местно на миометрии. Одним из важных утеротонинов является окситоцин, который играет особую роль в третьем периоде родов и после выделения последа, предотвращая развитие послеродового кровотечения. До сих пор неясно, как миометрии из поддерживающего состояния беременности ведет к началу родов при достижении 260 дней от момента имплантации плодного яйца т. е. момент наступления родов в определенной степени запрограммирован. Исследованиями Me Lean и соавт. (1995) по определению содержания в крови беременных плацентарного кортико-либерина установлено, что при сроке 18 нед. беременности можно выделить группы с нормальным, преждевременным и запоздалым началом родов. К началу родов содержание кортиколиберина резко возрастает и поэтому полагают, что это можно рассматривать как сигнал начала родов. В поддержании беременности и наступления родов большую роль придают плодово-материнской системе (Casey M.L и Mac Donald P.C., 1988). Предлагается три гипотезы начала родов: 1) снижение уровня прогестерона; 2) окситоциновая теория; 3) изменения плодово-материнской связи. Исследованиями G.C.Liggins (1973), J.R. Challis и D.M.Olson (1988) доказано, что у овец началу родов предшествует возрастание концентрации кортизола у плода и резкое снижение уровня прогестерона; затем отмечается подъем содержания эстрадиола — 17 р. У человека и приматов не установлено падения прогестерона в материнской плазме перед началом родов. Его снижение наблюдается после рождения плаценты и является сигналом для лактогенеза. Введение АКТГ или кортизола плоду человека не вызывает родов. Введение дексаметазона беременной приводит к значительному замедлению образования эстрогенов (ингибирует образование адреналового плодового C19-стероида). В наступлении родов имеют значения промежуточные модуляторы. Полагают, что при преждевременных родах нередко играет роль бактериальный токсин, который вызывает увеличение гидролиза глицерофосфолипида, подъем содержания свободной арахидоновой кислоты и образование простагландинов. Роль промежуточного модулятора в развитии родовой Деятельности играет интерлейкин-1 (ИЛ-1). Этот возможный интергормон-интермодулятор во многих тканях вызывает Усиленный гидролиз глицерофосфолипидов, способствует выделению арахидоновой кислоты и образованию простагландинов и факторов активации тромбоцитов. Синтез окситоцина осуществлен в 1953 г. Он синтезируется в супраоптичсеких и паравентрикулярных невронах и транспортируется по аксонам в невральной зоне задней доли гипофиза в мембранные везикулы, где хранится и затем выделяется. Окситоцин является очень сильным утеротоником и в небольших концентрациях вызывает сокращения сенсибилизированной матки. Новейшими исследованиями установлено, что роль окситоцина в вызывании родов обусловлена: 1) значительным возрастанием количсетва рецепторов окситоцина в миометрии в конце беременности и 2) воздействием окситоцина на выделение простагландинов. Новые данные в отношении физиологии и биохимии окситоцина доказывают его роль во втором периоде родов и после рождения плода, но не раньше. Нет доказательств того, что окситоцин играет роль в подготовке матки к родам. Инфузия окситоцина эффективна для вызывания родов при доношенной беременности и зрелой шейки матки, а также в первом периоде родов. Установлено, что увеличение количества рецепторов окситоцина в миометрии или в децидуальной оболочке при поздних сроках беременности может быть столь значительным, что гормон может быть эффективным даже при неизменном уровне в крови. T.Chard (1974), S. Kimura (1983), K.De Gust (1985) указывают на большое значение плодового окситоцина и вазоп-рессина в возникновении родовой деятельности. Установлено, что при концентрации окситоцина в околоплодных водах, превышающей 10 пг/мл, родовая деятельность начинается спонтанно. В конце беременности возрастает количество рецепторов к окситоцину и увеличивается чувствительность миомет-рия. Установлено, что концентрация окситоцина в плазме беременных женщин колеблется от 2 до 10 пг/мл (Leake R.D. et al., 1981). Концентрация окситоцина в крови не изменяется во время родов; исключением является конец родов. Инфузия окситоцина со скоростью 20 пг/мин обычно вызывает роды у женщин (1 ME окситоцина = 2 пг). Она эффективна, если его концентрация возрастает в 3-7 раз. Уровень окситоцина возрастает при кормлении грудью и способствует отделению молока. Окситоцин является синергистом с другими утеротониками такими как простагландин. Действие окситоцина на сократительную активность матки зависит от гормонального фона, прежде всего, от уровня эстрогенов и прогестерона, а также от функционального состояния матки, ее спонтанной активности. Нарастание концентрации окситоцина во время беременности, по-видимому, существенно влияет на подготовку матки к активной родовой деятельности. Механизм действия окситоцина на миометрии связан с уменьшением мембранного потенциала мышечной клетки. Известную роль в этом процессе играет транспорт ионов натрия и кальция. Окситоцин влияет на скорость связывания ацетилхолина рецепторами миометрия, участвует в механизме высвобождения ацетилхолина из его связанного состояния.Действие окситоцина проявляется за счет угнетения активности холинэстеразы, изменения электролитных свойств клеток, возбуждения а-адренорецепторов матки и других явлений. Действие окситоцина, стимулирующего сокращение всех отделов матки, наиболее выражено в процессе развившихся родов и направлено для их завершения и предотвращения послеродовых кровотечений. Количество окситоцина в организме зависит от активности разрушающего фермента — окситоциназы, вырабатываемого плацентой. В плазме крови окситоцин находится в динамическом равновесии, обусловленном активностью окситоциназы. С увеличением срока беременности наблюдаются повышение концентрации окситоцина и нарастание активности окситоциназы в крови и плаценте. Простагландины (ПГ) представляют собой ненасыщенные жирные кислоты с 20 углеродными атомами, окружающими скелет молекулы простаноевой кислоты. Различают четыре серии натуральных простагландинов: Е, F, А и В. Особый интерес в репродуктивной физиологии представляют соединения серии Е и F. Синтез простагландинов F2a и Е2 из ненасыщенных жирных кислот был осуществлен S. Bergstrom и соавт. (196-1) и D.A. Van Dorp et al. (1964), после чего эти вещества начали использоваться в клинике. Позже S. Bergstrom и соавт. Были удостоены Нобилевской премии за синтез простагландинов и фундаментальные исследования в этой области. В настоящее время доказана роль ПГ в наступлении родов. Установлено следующее: 1) уровень ПГ в амниотической жидкости, в материнской крови, моче и тканях матки в родах возрастают; 2) простагландины ПГF2a и ПГЕ2, введенные в любые сроки беременности приводят к сокращению миомет-рия и вызывают аборт или роды; 3) простагландины эффективны для вызывания родов при введении их per os, в амнио-тическую жидкость, внутривенно, экстраовулярно; 4) введение ингибиторов синтеза ПГ ведет к пролонгированию беременности и удлинению родового процесса; 5) введение ингибиторов синтеза ПГ эффективно при лечении преждевременных родов; 6) простагландины могут использоваться как утеротропины. Источником образования простагландинов является арахидоновая кислота. Знание механизма синтеза простагландинов в тканях является источником для понимания процесса родов. Биосинтез простагландинов осуществляется в различных тканях: простациклин ПГI2 синтезируется в миометрии, Е2 — в амнионе и хорионе, ПГF2a — в децидуальной ткани. Нет четких доказательств увеличения скорости образования ПГ во внутриматочных тканях до начала родов. Тогда как в родах имеется резкое возрастание концентрации ПГ Е2 и ПГF2a в амниотической жидкости. Возрастает также концентрация метаболитов ПГF2a, а именно 13-14 дегидро —15 кето ПГF2a в амниотической жидкости, крови и моче. С другой стороны нет четкого доказательства возрастания уровня ПГЕ2 (или метаболитов) в материнской крови (Mitchell M. D., 1988). ПГF2a может продуцироваться в децидуальной оболочке и в миометрии, но не в плодовых мембранах, однако возрастание концентрации ПГР2а и его метаболитов во время родов отмечено в амниотической жидкости, крови матери и моче. Важно подчеркнуть, что амниотическая жидкость способствует сохранению простагландинов. Так, период полураспада ПГF2a и Е2 в крови составляет 6-8 мин, тогда как в амниотической жидкости он колеблется от 4 до 6 часов. Существует гипотеза, что децидуальная активность синхронна с началом родов. Во время родов в амниотической жидкости аккумулируются биологически активные вещества, а именно, арахидоновая кислота, простагландины, активный тромбопласти-новый фактор, цитокины. Концентрация арахидоновой кислоты в амниотической жидкости в родах возрастает в 5-10 раз. Важную роль в возникновении родовой деятельности играют простагландины Е и F. Механизм действия их на сократительную деятельность матки изучен недостаточно. Полагают, что механизм стимулирующего действия на матку реализуется деполяризацией клеточных мембран и освобождением ионов кальция (Са2+), что ведет к активации киназой легкой цепи миозина, фосфорилированию миозина и взаимодействию фосфорилированного миозина и актина (Carsten M.E., Miller J.D., 1983), а возможно, их прямым стимулирующим влиянием на гипофиз, в результате усиливается синтез окси-тоцина (Gillespie A., 1973). Установлено, что при сочетанном применении ПГЕ2 или ПГF2a с окситоцином эффективность действия смеси выше, чем одного простагландина (Чернуха Е.А., 1983; BeazleyJ.M., 1979). Установлено, что манипуляции с плодными оболочками при вагинальном исследовании (отслаивание, введение баллона), амниотомия, манипуляции с шейкой матки способствуют выработке ПГР2а и его метаболитов (Mitchell M.D., 1976; Mortimer G. et al, 1985; Me Colgin S.W. et al., 1993). Роль плода в вызывании родов. Плодовый сигнал (триггер) начала родов может быть передан несколькими путями. Так у овцы сигналом к родам является подъем кортизола у плода и снижение содержания прогестерона. У человека несомненную роль в развитии родовой деятельности играет плод, его гипофизарно-надпочечниковая система, сигналы от трофобласта. Масса плода, генетическая завершенность развития, иммунные взаимоотношения плода и матери влияют на начало родовой деятельности и течение родов. Сигналы, поступающие из организма зрелого плода, обеспечивают информирование материнских компетентных систем, ведут к подавлению синтеза иммуносупрессорных факторов, в частности пролактина, а также хориального гонадотропина. Меняется реакция организма матери к плоду как трансплантату. В фетоплацентарном комплексе меняется стероидный баланс в сторону накопления эстрогенов, увеличивающих чувствительность адренорецепторов к норадреналину, окситоцину, простагландину. Установлено, что при аномалиях развития плода (анэнцефалия, микроцефалия, гидроцефалия, гипоплазия надпочечников) нарушается синтез эстрогенов у матери, так как они образуются из предшественников, вырабатываемых в надпочечниках плода (дегидроэпиандростерои и 16-оксидегидроэ-пиандростерон), нарушается синтез плодового окситоцина и выработка кортизола (Turnbull A.C., 1974; Nwosu N.C.et al., 1975; Turburn G.D., 1983). На сократительную функцию матки оказывает влияние внутриматочное давление, величина плода, что опосредуется раздражением хемо- и барорецепторов. Доказательств этого — частные преждевременные роды при многоводии и многоплодии. Особый интерес представляет физиология сокращения матки. Установлено, что финальным исходом начала маточного сокращения является возрастание внутриклеточной концентрации ионов кальция (Са2+) в миометрии гладкомышеч-ных клеток в ответ на действие утеротонина. Определенную роль играют "щелевые контакты" (gap junction) между клетками миометрия для обеспечения сокращения миометрия и течения родового процесса. Эти "щелевые контакты" проявляются при доношенной беременности и их количество увеличивается в родах (Garfield R.E.G., 1988; Ulstrom U., 1992), а затем они исчезают в течение 24 часов после родов. Эти "щелевые контакты" отсутствуют в миометрии при недоношенной беременности и наблюдаются при преждевременных родах и при родах спонтанных или индуцированных. В исследованиях на животных in vivo и in vitro установлено, что прогестерон предотвращает, а эстрогены способствуют образованию связей ("щелевых контактов"), и для этого необходим синтез протеина, важна также роль простагландинов, тромбоксана и, возможно, эндоперекиеи. Угнетение синтеза ПГ подавляет образование "щелевых контактов". Сокращения гладкомышечных волокон миометрия отличаются от такового скелетной мускулатуры. Расположение волокон в миометрии в различном направлении способствует усилению выталкивающей силы матки независимо от положения и предлежания плода. Молекулярная регуляция гладкомышечного сокращения. В мышечном сокращении основную роль играет миозин. У молекулы миозина есть функциональные "головная" и "хвостовая" части. Шаровидная (глобулярная) "головная" часть является: местом взаимодействия с актином, т.е. местом, где генерируется сила сокращения; местом, где находится АТФ-азы, под действием которой гидролизуется АТФ, и, тем самым, химическая энергия превращается в физическую силу; местом, где расположены легкие цепи миозина с низкой молекулярной массой (=20 000), фосфорилирование которых представляет собой ключевую реакцию в регуляции сокращения, т.е. во взаимодействии актина и миозина в гладкой мышце. В связи с этим важно помнить, что для генерации мышечного сокращения необходимо поступление ионов кальция (Са2+) из внеклеточных или внутриклеточных запасов. Са2+ накапливается внутри клеток в пузырьках саркоплазматического ретикулума. В гладких мышцах взаимодействие актина и миозина регулируется ферментативными фосфорилированием (или дефосфорилированием) легких цепей миозина (Stull J.T. et al., 1980). В клетках миометрия взаимодействие актина с миозином происходит только после фосфорилирования легкой цепи миозина. Это фосфорилирование катализируется ферментом киназа легкой цепи миозина; важно, что этот фермент активизируется Са2+. Соединения, которые увеличивают содержание внутриклеточного свободного Са2+, усиливают сокращения гладкой мускулатуры миометрия. Кальций связывается с кальмодулином (регуляторным белком, который опосредует эффект кальция), который в свою очередь, связывается с киназой легкой цепи миозина и активирует ее. Ситуации, уменьшающие уровень свободного внутриклеточного кальция, способствуют расслаблению. Дефосфорилирование легкой цепи миозина под действием фосфотаз также способствует расслаблению мышцы. Актин не взаимодействует с нефосфорилизированным миозином. Соединения, которые повышают внутриклеточную концентрацию циклического АМФ (цАМФ), усиливают релаксацию матки. Эти соединения включают b-адренергические агонисты. Считает ся, что цАМФ вызывает снижение внутриклеточного Са2+, хотя это не доказано. Согласно другой точке зрения, происходит инактивация фермента киназа легкой цепи миозина в результате его фосфорилирования под воздействием цАМФ-зависимого фермента протеин-киназа. Снижение активности фосфорилирования фермента объясняется снижением его сродства к кальмодулину; для проявления активности этого фермента его взаимодействие с кальмодулином должно быть обязательно. Можно считать, что регуляция сократительной активности миометрия на клеточном уровне осуществляется при посредстве киназы легкой цепи миозина, на активность которой влияет кальций. Одновременно следует учитывать и дефосфо-рилирование её под действием соответствующей фосфотазы. Активация сокращения осуществляется путем взаимодействия фосфорилированного миозина и актина с образованием фосфорилированного актомиозина. Акт родов протекает при наличии сформированной родовой доминанты, объединяющей в динамическую систему как высшие нервные центры, так и исполнительные органы. В формировании родовой доминанты большое значение имеет воздействие половых гормонов на различные образования центральной и периферической нервной системы. Центральная нервная система осуществляет высшую и тонкую регуляцию родового акта. Значительное возрастание электрической активности головного мозга наступает за 1-1,5 нед. до наступления родов. Большое значение для возникновения родовой деятельности и правильного ее течения имеет подготовленность женского организма, готовность матки, а также чувствительность миометрия к воздействию контрактильных веществ. Дата добавления: 2016-06-06 | Просмотры: 978 | Нарушение авторских прав |