|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Вторые нейроны
После того как афферентные волокна входят в спинной мозг, они разделяются в соответствии с размерами, при этом толстые миелинизирован-ные волокна располагаются медиально, а тонкие немиелинизированные — латерально. Перед переключением на вторые нейроны в сером веществе ипсилатеральных задних рогов спинного мозга, некоторые волокна болевой чувствительности могут в составе проводящего пути Лиссауэра подниматься или опускаться на один-три сегмента спинного мозга. Во многих случаях связующим звеном между первыми и вторыми нейронами являются вставочные нейроны. Согласно Рекседу, серое вещество спинного мозга разделено на 10 пластин, или слоев, идущих в направлении от продолговатого мозга каудально (рис. 18-2 и табл. 18-4). Первые шесть пластин, которые формируют задние рога, воспринимают всю афферентную информацию и являются основным местом модуляции боли посредством восходящих и нисходящих влияний (см. далее). Вторые нейроны функционально представляют собой либо ноци-цептивно-специфичные нейроны, либо нейроны широкого динамизма. Ноцицептивно-специфичные нейроны реагируют только на ноцицептивные стимулы, в то время как нейроны широкого динамизма воспринимают по волокнам Аβ, Аδ и С также и не-ноцицептивные афферентные импульсы.
Рис. 18-1. Пути проведения болевой чувствительности
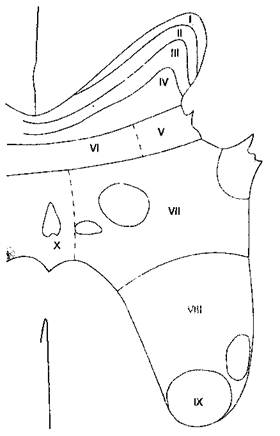
Рис. 18-2. Слои серого вещества спинного мозга по Рекседу Ноцицептивно-специфичные нейроны находятся в I пластине, имеют дискретные соматические рецепторные поля, в норме находятся в состоянии покоя и реагируют только на высокопороговую ноцицептивную стимуляцию, плохо кодируя ее интенсивность. Нейроны широкого динамизма — наиболее распространенный тип клеток в веществе задних рогов. Хотя нейроны широкого динамизма находятся во всех участках задних рогов, в большей степени они сконцентрированы в V пластине. При повторяющейся стимуляции частота импульсации нейронов широкого динамизма ступенчато возрастает по экспоненте ("реакция испуга"), даже если интенсивность стимула не увеличивается. Подобно ноцицептивно-специфичным нейронам, нейроны широкого динамизма имеют обширные рецепторные поля. Большинство ноцицептивных волокон типа С имеют коллатерали или заканчиваются на вторых нейронах в I, II и, в меньшей степени, в V пластине. В отличие от них, ноцицептивные волокна типа Аδ образуют синапсы преимущественно в пластинах I и V, и, в меньшей степени, в X пластине. Клетки I пластины реагируют главным образом на ноцицептивные импульсы из кожи и глубоких соматических структур. Пластина II, называемая также студенистым веществом (substantia gelatinosa), содержит много вставочных нейронов и играет большую роль в восприятии и модуляции ноцицептивных импульсов с кожи. Пластина II представляет особый интерес, поскольку считается, что именно она — главное место действия опиоидов. Пластины III и IV воспринимают в основном неноцицептив-ные сенсорные импульсы. Передние (двигательные) рога формируются преимущественно VIII и IX пластинами. VII пластина носит название латерального промежуточного столба и содержит тела преганглионарных симпатических нейронов. Висцеральные афферентные волокна заканчиваются в основном в V пластине, в меньшей степени — в I. Эти две пластины представляют собой зону центральной конвергенции соматических и висцеральных импульсов. V пластина реагирует на ноцицептивные и неноцицептивные сенсорные импульсы, воспринимает соматическую и висцеральную боль. Феномен конвергенции (схождения) висцеральных и соматических сенсорных импульсов клинически проявляется иррадиацией боли (табл. 18-3). В отличие от соматических волокон, висцеральных ноцицептивных волокон меньше, они более диффузно распределены, активируют относительно большее количество нейронов в спинном мозге и не имеют соматотопической организации. А. Спиноталамический путь. Аксоны большинства вторых нейронов направляются на противоположную сторону спинного мозга через переднюю белую спайку, включаются в состав спи-ноталамического пути и достигают таламуса, ретикулярной формации, ядра шва и центрального серого вещества (околоводопроводного). Спино-таламический путь — главный путь проведения болевой чувствительности — проходит в переднела-теральном отделе белого вещества спинного мозга. Этот восходящий путь можно разделить на латеральный pi медиальный. Аксоны второго нейрона латерального спиноталамического пути (синоним — неоспиноталамический путь) заканчиваются синапсом на клетках вентральных заднелатераль-ных ядер таламуса. Латеральный спиноталамичес-кий путь обеспечивает восприятие дискримина-тивных аспектов боли — таких как локализация, интенсивность и длительность. Аксоны второго нейрона медиального спиноталамического пути (синоним — палеоспиноталамический путь) заканчиваются синапсом на клетках медиальных отделов таламуса. Медиальный спиноталамический путь обеспечивает восприятие вегетативных и эмоциональных аспектов боли. Некоторые волокна спиноталамического тракта заканчиваются на клетках центрального серого вещества и могут служить важным связующим звеном между восходящими и нисходящими проводящими путями (см. ниже). Коллатеральные волокна образуют синапсы на нейронах активирующей ретикулярной формации и гипоталамуса, что обеспечивает реакцию активации при болевой стимуляции. ТАБЛИЦА 18-4. Пластины (слои) серого вещества спинного мозга
Б. Альтернативные пути проведения болевой чувствительности. Аналогично путям проведения эпикритической чувствительности, ноцицептивные волокна могут проходить диффузно, ипсила-терально и контралатералъно, поэтому некоторые больные продолжают ощущать боль после хирургического пересечения контралатерального спиноталамического пути. Таким образом, помимо спиноталамического пути, играют весьма важное значение и другие восходящие пути проведения болевой чувствительности. Считают, что спиноре-тикулярный путь опосредует реакцию активации и реакцию вегетативной нервной системы на боль. Спиномезенцефалический путь может активировать нисходящие антиноцицептивные влияния, потому что некоторые его волокна заканчиваются на клетках центрального серого вещества. Спино-гипоталамический и спинотеленцефалический пути активируют гипоталамус и влияют на эмоционально-поведенческие аспекты восприятия боли. Спинно-шейный путь, не пересекая среднюю линию, поднимается до латерального шейного ядра, которое отдает волокна к контралатеральному та-ламусу; этот путь, вероятно, является главным альтернативным путем проведения болевой чувствительности. Наконец, некоторые волокна в задних столбах серого вещества (которые обеспечивают перцепцию тактильной и проприоцептивной чувствительности) могут проводить болевые импульсы; этот путь в спинном мозге проходит медиально и ипсилатерально. В. Интеграция с симпатической и двигательной системами. Соматические и висцеральные афферентные волокна образуют многочисленные синапсы с двигательными и симпатическими волокнами в спинном и продолговатом мозге, а также в высших мозговых центрах. Афферентные нейроны задних рогов переключаются на двигательные нейроны передних рогов с помощью прямого синаптического контакта, а также через вставочный нейрон. Эти синапсы обеспечивают рефлекторную мышечную активность (как физиологическую, так и патологическую) при болевом раздражении. Контакты между афферентными ноцицептивными нейронами и симпатическими нейронами в латеральном промежуточном столбе обеспечивают вегетативную реакцию на боль — вазоконстрикцию, спазм гладкой мускулатуры, выброс катехоламинов локально и из мозгового слоя надпочечников. Дата добавления: 2015-02-06 | Просмотры: 1256 | Нарушение авторских прав |