|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
А. Усиление боли (облегчение восприятия)В спинном мозге существуют по крайней мере три механизма центральной сенситизации: 1. "Реакция испуга" (см. выше) и сенситизация вторых нейронов пути болевой чувствительности. При повторяющейся стимуляции частота импульсации нейронов широкого динамизма ступенчато возрастает, даже если интенсивность повторного стимула не увеличивается. Кроме того, для нейронов широкого динамизма характерна спонтанная активация даже после прекращения поступления импульсов по афферентным волокнам типа С. 2. Расширение рецепторных полей. Вторые афферентные нейроны, расположенные в задних рогах, расширяют свои рецепторные поля таким образом, что окружающие их нейроны отвечают на те стимулы, к которым были до того интактны. 3. Гипервозбудимость сгибательных рефлексов. Усиление сгибательных рефлексов наблюдается как ипсилатерально, так и контралате-рально.
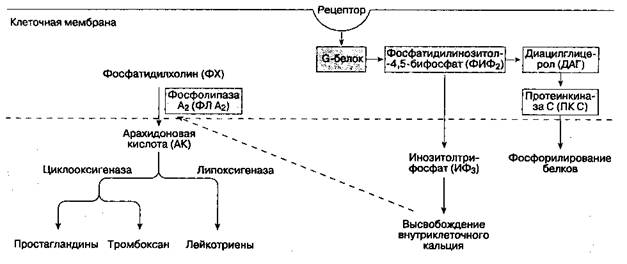
Рис. 18-3. Фосфолипаза С (ФЛ С) катализирует гидролиз фосфатидилинозитола-4,5-бифосфата (ФИФ2), при этом образуются инозитолтрифосфат (ИФ3)и диацилглицерол (ДАГ). Важную роль играет протеинкиназа С (ПК С). Фосфолипаза A2 (ФЛ A2) катализирует превращение фосфатидилхолина (ФХ) в арахидоновую кислоту (AK) К нейромедиаторам центральной сенситизации относятся субстанция P, пептид, ассоциированный с геном кальцитонина, вазоинтестинальный пептид, холецистокинин, ангиотензин, аланин, а также возбудительные аминокислоты — L-глютамат и L-аспартат. На мембране нейронов находятся рецепторы, связанные с G-белком. Нейромедиаторы взаимодействуют с этими рецепторами, что изменяет возбудимость мембраны нейрона. Тонкие механизмы этого взаимодействия включают активацию внутриклеточных вторичных мессенджеров, фосфорилирование белков, высвобождение ионов кальция из внутриклеточных депо (см. рис. 18-3). Активируя NMDA- и нeNMDА-рецепторные механизмы, глютамат и аспартат играют важную роль в процессе "реакции испуга" (NMDA — это N-метил D-аспартат). Полагают, что эти аминокислоты в значительной степени ответственны за индукцию и поддержание центральной сенситизации. Активация NMDA-рецепторов увеличивает концентрацию внутриклеточного кальция в спинномозговых нейронах и активизирует фосфолипазу С (ФЛ С). Возросшая концентрация внутриклеточного кальция приводит к активации фосфолипазы A2 (ФЛ A2), которая катализирует превращение фосфатидилхо-лина (ФХ) в арахидоновую кислоту (AK), из которой, в свою очередь, образуются простагландины. Фосфолипаза С (ФЛ С) катализирует гидролиз фосфатидилинозитола-4,5-бифосфата (ФИФ2) на инозитолтрифосфат (ИФ3) и диацилглицерол (ДАГ), которые являются вторичными мессендже-рами. ДАГ активирует протеинкиназу С (ПК С). Активация NMDA-рецепторов, кроме того, индуцирует синтетазу оксида азота и, следовательно, способствует образованию этого соединения. Простагландины и оксид азота облегчают высвобождение возбудительных аминокислот в спинном мозге. Следовательно, такие ингибиторы циклооксигена-зы, как ацетилсалициловая кислота и нестероидные противовоспалительные препараты, дают важный аналгегпический эффект на уровне спинного мозга. Б. Ослабление боли (ингибирование, подавление болевой чувствительности). Проведение ноци-цептивного импульса в спинном мозге может быть ингибировано сегментарной активностью непосредственно на уровне спинного мозга, а также нисходящими влияниями из супраспинальных центров. 1. Сегментарное ингибирование. Активация крупных афферентных волокон, опосредующих эпикритическую (неноцицептивную) чувствительность, ингибирует активность нейронов широкого динамизма и проведение импульсов по спинотала-мическому пути. Кроме того, ноцицептивная стимуляция в одной области ингибирует активность нейронов широкого динамизма и, соответственно, боль в другой области тела. Эти наблюдения подкрепляют "шлюзовую" теорию распространения болевых импульсов в спинном мозге. Глицин и γ-аминомасляная кислота (ГАМК) -это аминокислоты, которые являются тормозными нейромедиаторами. Они играют важную роль в сег-ментарном ингибировании болевых ощущений на уровне спинного мозга. Антагонисты глицина и γ-аминомасляной кислоты вызывают выраженную активацию нейронов широкого динамизма, что приводит к аллодинии и гиперестезии. Существуют два подтипа ГАМК-рецепторов: ГАМКА, агонистом которых является мусцимол, и ГАМКВ, агонистом которых служит баклофен. Сегментарное ингибирование опосредовано через ГАМКв-рецепторы, активация которых ведет к повышению проницаемости клеточной мембраны для ионов калия. ГАМКА-ре-цепторы функционируют как канал для ионов хлора, их активация увеличивает проницаемость мембраны для Сl-. Бензодиазепины потенцируют влияние аго-нистов на ГАМКА-рецепторы. Активация глициновых рецепторов также увеличивает проницаемость мембраны нервной клетки для ионов хлора. Стрихнин и столбнячный токсин являются антагонистами глициновых рецепторов. Кроме того, глицин стимулирует NMDA-рецепторы, поэтому эффекты, которые он дает, сложнее эффектов ГАМК. Аденозин тоже модулирует ноцицептивную активность в задних рогах спинного мозга. Известны по меньшей мере два рецептора: A1, активация которого ингибирует аденилатциклазу, и A2, стимуляция которого, напротив, ее активирует. Антиноцицептив-ное действие аденозина проявляется через A1-рецепторы. Метилксантины, ингибируя фосфодиэстеразу, блокируют антиноцицептивный эффект аденозина. 2. Супраспинальное ингибирование. Некоторые супраспинальные структуры посылают волокна в спинной мозг, ингибируя передачу болевых импульсов в задних рогах. Эти структуры включают центральное серое вещество (оно окружает водопровод мозга), ретикулярную формацию и ядро шва. Стимуляция центрального серого вещества вызывает у человека аналгезию всего тела. Аксоны этих нисходящих путей оказывают ингибирующее влияние — как пресинаптическое (на первые нейроны путей проведения болевой чувствительности), так и постсинаптическое (на вторые и вставочные нейроны). Антиноцицептивное действие опосредовано через α2-адренорецепторы, серотониновые рецепторы, а также μ-, δ- и χ-опиатные рецепторы. Роль моноаминов в ингибировании боли объясняет аналгети-ческий эффект антидепрессантов, которые блокируют обратный захват катехоламинов и серотонина. Стимуляция этих рецепторов (которые также связаны с G-белками) активирует вторичные мессендже-ры, что открывает калиевые каналы и ингибирует высвобождение кальция из внеклеточных депо. Антиноцицептивные нисходящие адренергичес-кие пути начинаются главным образом в сером веществе коры и ретикулярной формации. Норадре-налин опосредует антиноцицептивный эффект через активацию пре- и постсинаптических α2-ад-ренорецепторов. По крайней мере часть аксонов из серого вещества коры переключаются на нейронах ядра шва и ретикулярной формации продолговатого мозга; из ядра шва серотонинергические волокна следуют в составе дорсолатерального канатика к нейронам задних рогов спинного мозга и ингибируют их активность. Эндогенная опиатная система (представленная главным образом ядром шва и ретикулярной формацией) действует через метионин-энкефалин, лейцин-энкефалин и β-эндорфин, антагонистом которых является налоксон. Эти опиоиды оказывают пресинаптическое действие, вызывая гипер-поляризацию первых нейронов и ингибируя высвобождение субстанции P; они также обеспечивают некоторое постсинаптическое ингибирова-ние. В противоположность им экзогенные опиоиды. оказывают главным образом постсинаптическое действие, ингибируя вторые или вставочные нейроны в студенистом веществе. Дата добавления: 2015-02-06 | Просмотры: 1367 | Нарушение авторских прав |