|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
ГЛЮКОНЕОГЕНЕЗ
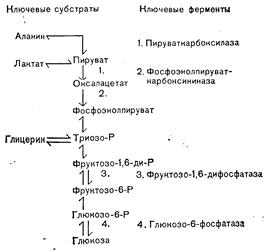
Этим термином называют образование глюкозы из неуглеводных источников. Основными субстратами-предшественниками, из которых могла бы образоваться глюкоза, являются пируват, лактат, глицерин, жирные кислоты с нечетным числом углеродных атомов и аминокислоты. Что касается последних, то все входящие в состав тканевых белков аминокислоты, за исключением лейцина, в конце концов действительно могут превращаться в глюкозу. Однако поглощение печенью аминокислот таково, что главным глюкогенным субстратом, высвобождаемым из периферических белковых запасов, является аланин. Жирные кислоты с четным числом углеродных атомов (составляющие более 95% от общего содержания жирных кислот) в печени млекопитающих не могут превращаться в глюкозу из-за отсутствия ферментов, необходимых для синтеза 4-углеродных дикарбоновых кислот из ацетил-СоА de novo. За исключением глицерина любые предшественники глюконеогенеза прежде чем превратиться в глюкозу должны превратиться в пируват и/или оксалацетат. Ферментативные стадии образования глюкозы из пирувата отличаются от тех, из которых состоит гликолиз, по трем пунктам, в которых происходят термодинамически необратимые реакции (рис. 10—4): 1) дефосфорилирование фосфоенолпирувата с образованием пирувата; 2) фосфорилирование фруктозо-1-фосфата с образованием фруктозо-1,6-дифосфата; 3) фосфорилирование глюкозы с образованием глюкозо-6-фосфата. Биологическая обратимость достигается с помощью четырех ферментов, принимающих участие только в глюконеогенезе: пируваткарбоксилазы, фосфоэнолпируваткарбоксикиназы; фруктозо-1,6-дифосфатазы и глюкозо-6-фосфатазы. Реакции, катализируемые этими ферментами, — ключевые регуляторные этапы глюконеогенеза, и протекают они в печени, почках и эпителии кишечника, но не в мышцах или в сердце. С количественной стороны наиболее важным местом глюконеогенеза в таких физиологических условиях, как голодание или физическая нагрузка, и при таких патологических состояниях, как диабет, является печень. Почки приобретают значение в качестве органа глюконеогенеза только при очень длительном голодании [9].
Рис. 10 — 4. Глюконеогенез. Основные субстраты глюконеогенеза и ключевые ограничивающие скорость ферментативные стадии. Постоянная регуляция глюконеогенеза зависит от присутствия субстратов, активности ферментов и гормональной среды. В состоянии натощак (после ночного голодания) и при кратковременном голодании снижение уровня инсулина в сыворотке способствует увеличению распада белка и мобилизации аминокислот, что обеспечивает снабжение процесса большим количеством предшественников глюкозы. В этих условиях мобилизуются и жирные кислоты, служащие главным субстратом окисления в печени. Вследствие этого повышается внутрипеченочный уровень ацетил-СоА, что приводит к активации ключевого фермента глюконеогенеза пируваткарбоксилазы, которая аллостерически активируется ацетил-СоА. «Чистым» результатом является увеличение как уровня субстратов, так и активности ферментов, необходимых для глюконеогенеза. Однако по мере продолжения голодания в течение длительных периодов ограничивающим скорость процессом становится доступность субстрата-предшественника, так как высвобождение аланина периферическими тканями существенно уменьшается,[10]. В отличие от голодания при большом количестве углеводов (например, в состоянии сытости) интенсивность глюконеогенеза снижается. Мобилизуется меньше жира прежде всего из-за того, что повышенная секреция инсулина, вызванная углеводами, тормозит высвобождение жирных кислот из депо (см. далее). В результате образуется меньшее количество ацетил-СоА и активность пируваткарбоксилазы снижается. Кроме того, в меньшей степени мобилизуются аминокислоты. В силу этих изменений поглощение печенью и превращение в глюкозу предшественников глюконеогенеза, особенно аланина, угнетается. Регуляторные воздействия таких гормонов, как глюкокортикоиды и глюкагон, на глюконеогенез реализуются также на этапах превращения пирувата в фосфоенолпируват (см. рис. 10—4). При этом глюкокортикоиды влияют на глюконеогенез еще в силу своего катаболического действия на тканевые белки, что увеличивает количество аминокислот-предшественников. Дата добавления: 2015-02-05 | Просмотры: 981 | Нарушение авторских прав |