|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
ДЕЙСТВИЕХотя глюкагон был обнаружен почти 60 лет назад в качестве гипергликемизирующего фактора, присутствующего в неочищенных препаратах инсулина, его физиологическая роль выяснилась лишь недавно. Ранние исследования наталкивались на невозможность воспроизведения чистого глюкагонодефицитного состояния без одновременного создания дефицита инсулина, а также на отсутствие данных об уровне в крови и скорости секреции глюкагона. Разработка радиоиммунологических методов и открытие соматостатина способствовали выяснению роли глюкагона в гомеостазе глюкозы. Физиологические приросты содержания глюкагона вызывают повышение уровня глюкозы в крови за счет стимуляции гликогенолиза и глюконеогенеза в печени (рис. 10—22). Наоборот, снижение концентрации глюкагона ниже исходного уровня приводит к снижению в печени продукции глюкозы [86]. Если прирост уровня глюкагона сопровождается небольшим (на 50—150%) приростом уровня инсулина в крови, продукция глюкозы в печени остается неизменной. Последний феномен определяет значение бигормональной реакции (повышение секреции как глюкагона, так и инсулина) на прием белковой пищи (рис. 10—23). Реакция инсулина, вызываемая белковой пищей, обеспечивает поглощение и утилизацию клетками содержащихся в ней аминокислот. Однако само по себе повышение уровня инсулина должно было бы снизить выход глюкозы из печени и тем самым вызвать гипогликемию. Одновременный же прирост уровня глюкагона препятствует проявлению такого эффекта инсулина и обеспечивает сохранение продукции глюкозы на стабильном уровне (см. рис. 10—23). Поскольку при приеме смешанной пищи не изменяется содержание глюкагона (см. рис. 10—13), можно предположить, что глюкагон в ходе эволюции приобрел роль регулятора гликемии главным образом при потреблении мяса. В отличие от инсулина, влияние которого на различные ткани-мишени продолжается до тех пор, пока не возникнет гипогликемия, глюкагон оказывает лишь кратковременное действие на печеночную продукцию глюкозы, продолжающееся всего 30—
Рис. 10—22. Стимулирующее влияние глюкагона на продукцию глюкозы органами брюшной полости (печенью) у здорового человека. Физиологическая гиперглюкагонемия вызывает быстрое повышение печеночной продукции глюкозы. Однако этот эффект является кратковременным, сохраняясь в течение менее 45 мин. Стимулирующее действие глюкагона на продукцию глюкозы, таким образом, определяется не абсолютной его концентрацией, а изменениями последней (по Felig P. et al., Diabetes, 1976, 25, 1901). 1 — глюкоза: 2 — инсулин. 60 мин (см. рис. 10—22) [84]. Однако, если по достижении данного уровня глюкагона в плазме произойдет его дальнейшее повышение или снижение, то соответственно повысится или снизится продукция глюкозы. Таким образом, глюкагон влияет на метаболизм глюкозы в печени путем изменения своего уровня, а не абсолютной концентрацией. Следовательно, печень сохраняет чувствительность к изменениям уровня глюкагона в крови, но эффект любого данного изменения этого уровня оказывается кратковременным, продолжаясь обычно 60 мин или менее [87].
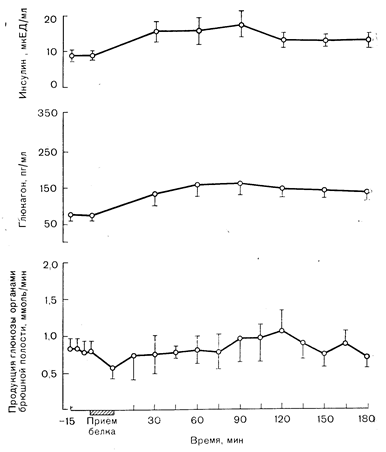
Рис. 10—23. Влияние приема белковой пищи на уровень инсулина и глюкагона в плазме, а также на спланхническую (печеночную) продукцию глюкозы у здорового человека. Повышение содержания глюкагона, вызываемое белковой пищей, необходимо для поглощения и анаболической утилизации содержащихся в ней аминокислот (см. рис. 19—9) (по данным Wharen J., Felig P., Hagenfeldt L. J., Clin. Invest., 1976, 58, 761).
Учитывая противоположность эффектов инсулина и глюкагона на печень, а также тот факт, что глюкоза подавляет секрецию глюкагона, но стимулирует секрецию инсулина, Unger и соавт. постулировали, что общий углеводный гомеостаз управляется не концентрацией каждого из этих гормонов порознь, а молярным соотношением инсулин/глюкагон (И/Г) [88, 89]. Применимость датой гипотезы к целому организму была поставлена под серьезное сомнение Sherwin и соавт., проводившими исследования с инфузией физиологических доз глюкагона. Эти исследования показали, что повышение глюкагона в плазме до уровня 300— 400 пг/мл (сопоставимого с наблюдающимся при различных гиперглюкагонемических состояниях) не изменяет толерантности к глюкозе у здоровых лиц. Таким образом, несмотря на выраженное уменьшение соотношения И/Г (с 34:1 до 6:1), изменений толерантности к глюкозе не появляется [84]. Это означает, что глюкагон может нарушать глюкозотолерантность только либо в условиях абсолютной недостаточности инсулина, либо при введении его в фармакологических дозах, либо, наконец, при глюкагонпродуцирующей опухоли (глюкагонома). Кроме того, хотя введение глюкозы в норме подавляет секрецию глюкагона, но пока секреция инсулина остается в пределах нормы, сохранение нормальной скорости исчезновения глюкозы из крови не требует гипоглюкагонемии [90, 91]. Помимо влияния на углеводный обмен, глюкагон участвует и в регуляции кетогенеза. В печени крыс, получавших физиологические дозы глюкагона, повышается продукция кетоновых тел из жирокислотных предшественников [15]. In vivo гиперкетонемия может встречаться и при отсутствии глюкагона, как, например, у больных с удаленной поджелудочной железой [81]. Однако у больных инсулинодефицитным диабетом гиперглюкагонемия еще больше увеличивает кетогенную способность печени (см. рис. 10—8). Эффекты глюкагона, заключающиеся в повышении уровня сахара в крови и кетоза, целиком определяются его действием на печень. При физиологических приростах уровня глюкагона не изменяются ни периферическое (внепеченочное) поглощение глюкозы, ни липолиз. Первый этап действия глюкагона на печень это — его связывание с рецепторами клеточной мембраны. Пока не ясно, однако, является ли рецепторный уровень существенным для модуляции действия этого гормона. Вторым медиатором эффекта глюкагона служит повышение содержания цАМФ. Это происходит путем стимуляции гормончувствительной аденилатциклазы в плазматической мембране. Повышение уровня цАМФ «запускает» реакцию фосфорилирования, приводящего к повышению активности ключевого фермента гликогенолиза — фосфорилазы и одновременному торможению гликогенсинтетазы (см. рис. 10—3). Что касается глюконеогенеза, то чувствительный к глюкагону этап этого процесса расположен на уровне превращения пирувата в фосфоенолпируват (см. рис. 10—4). Введение глюкагона снижает уровень калия и кальция в сыворотке крови, секрецию соляной кислоты и ферментов поджелудочной железы, повышает секрецию гормона роста и увеличивает сократимость миокарда. Однако все эти эффекты требуют фармакологических доз гормона.
Дата добавления: 2015-02-05 | Просмотры: 1011 | Нарушение авторских прав |