|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
НАСЫЩЕННОСТЬ РЕЦЕПТОРОВ И АКТИВАЦИЯ РЕАКЦИЙ КЛЕТОК-МИШЕНЕЙБольшинство исследований, посвященных результатам гормонрецепторных взаимодействий, свидетельствуют о том, что активация аденилатциклазы и реакций клеток-мишеней является функцией насыщенности рецепторов. Во многих тканях, однако, сродство связывания рецепторов оказывается меньшим, чем концентрация гормона требуемая для развития полумаксимальной биологической реакции, а полная биологическая реакция называется насыщением небольшой части имеющихся рецепторов. Даже в тех случаях, когда все большее насыщение рецепторов сопровождается ростом реакций в широких пределах связывания гормона, степень реакции обычно непропорциональна числу занятых рецепторов. Кроме того, известно, что многие гормоны способны стимулировать продукцию гораздо большего количества цАМФ, чем это необходимо для индукции максимальной биологической реакции в клетке-мишени. Такие наблюдения привели к выводу, что многие гормончувствительные ткани характеризуются «запасом» или «избытком» рецепторов, или «рецепторным резервом», что аналогично ситуации, описанной Stephenson при изучении тканей, чувствительных к лекарственным веществам [14]. Несомненно, что такие рецепторные участки клетки не являются лишними; предполагают, что они играют роль резервуара поверхностных рецепторов, обеспечивающего возможность повышения чувствительности клеток-мишеней к активирующему влиянию низких концентраций гормона. Таким образом, высокая плотность поверхностных рецепторов могла бы способствовать быстрой ассоциации и достижению критических или пороговых уровней насыщенности, обусловливающих активацию клетки-мишени.
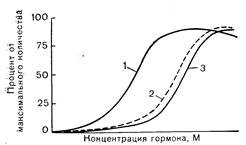
Рис. 4—5. Связывание гормона, образование цАМФ и продукция стероидов при стимуляции клеток половых желез и надпочечников тропным гормоном. 1 — реакция (например, продукция стероида); 2 — связанный гормон: 3 — цАМФ.
Важно подчеркнуть, что термин «запас» применяется в относительном смысле и что степень избыточности рецепторов может варьировать в зависимости от регистрируемой биологической реакции. Например, в отношении активации аденилатциклазы редко наблюдается «запас» рецепторов, поскольку существует четкая корреляция между связыванием и активностью аденилатциклазы, хотя иногда может встречаться и нелинейная зависимость между насыщенностью рецепторов и реакцией фермента. Подобно этому, связывание гормона с интактными клетками-мишенями обычно сопровождается прогрессирующим увеличением продукции цАМФ, и большинство тканей не содержит избытка рецепторов, который не был бы сопряжен с аденилатциклазой. Однако при регистрации более «дистальных» реакций, таких, как мышечное сокращение, стероидогенез, окисление глюкозы, липолиз и транспорт ионов, часто обнаруживается, что максимальная биологическая реакция воспроизводится при насыщении только небольшой фракции популяции рецепторов (рис. 4—5). Совершенно очевидно, что относительно большой пул рецепторов в мембране клетки-мишени должен облегчать активацию малого их числа (необходимого для достижения порога индукции клеточной реакции). Избыток рецепторов легче всего установить путем непосредственного изучения связывания меченых гормонов с интактными плетками- и тканями-мишенями. Это особенно заметно в интерстициальных клетках семенников, где достаточно насытить менее 1% общей популяции рецепторов, чтобы вызвать полную активацию стероидогенеза и продукции тестостерона [15]. Однако в тех же самых клетках дальнейшее связывание меченого гонадотропина сопровождается прогрессированием роста продукции и высвобождения цАМФ. Это свидетельствует о том, что большинство или даже все связывающие места, выявляемые при взаимодействии с радиоактивным гормоном, являются функционально активными рецепторами, способными стимулировать активность аденилатциклазы и образованием цАМФ. В некоторых тканях связывание гормона и последующая биологическая реакция тесно сопряжены во всем диапазоне насыщенности рецепторов. Так, насыщенность рецепторов ангиотензина II в клубочковой зоне коры надпочечников коррелирует с увеличением стероидогенеза и продукции альдостерона во всем диапазоне связывания гормона. Пусть даже зависимость между связыванием ангиотензина II и продукцией альдостерона и нелинейна, но такая корреляция указывает на отсутствие «запаса» рецепторов в отношении стероидогенеза в клетках этого типа [16]. Подобно этому, на изолированных клетках вилочковой железы наблюдали тесную корреляцию между связыванием инсулина и стимуляцией транспорта аминокислот. Возможно, существуют широкие колебания зависимости между насыщенностью рецепторов и специфической реакцией клеток в разных тканях-мишенях от одной крайности в случае интерстициальных клеток семенников, полная активация которых происходит при минимальной насыщенности рецепторов, до другой — почти непрерывной зависимости между насыщенностью и реакциями, что встречается в клетках клубочковой зоны коры надпочечников и вилочковой железы. В клетках одного типа, например жировых клетках, повышение степени насыщения рецепторов при воздействии инсулина вызывает последовательный ряд реакций. Угнетение липолиза в этом случае происходит при очень низкой насыщенности рецепторов, стимуляция обмена глюкозы оказывается максимальной при насыщении всего 2—3% рецепторов, а стимуляция транспорта аминокислот и синтеза белка наблюдается при высокой степени насыщенности рецепторов [17]. Как правило, в тех случаях, когда связывание гормона и биологические реакции тесно коррелируют между собой, именно рецепторы лимитируют активацию клеток-мишеней. Если же имеется «запас» рецепторов, лимитирующими являются более «дистальные» элементы; лимитирующий этап при этом в случае реакций, опосредуемых цАМФ, локализуется, по-видимому, на уровне активации протеинкиназ. Дата добавления: 2015-02-05 | Просмотры: 1097 | Нарушение авторских прав |