|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
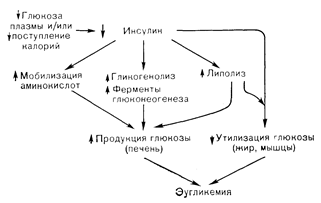
ГОЛОДАНИЕ ЗДОРОВЫХ ЛИЦВ норме человек принимает пищу с перерывами, которые варьируют от 2 до 14 ч. Иногда, однако, либо в силу добровольного отказа ют пищи, либо из-за какого-то привходящего заболевания, либо, наконец, из-за отсутствия пищи людям приходится голодать в течение более длительного времени. Метаболическая реакция на голодание объединяет в себе гормональные и субстратные изменения, направленные на: 1) сохранение продукции глюкозы (гликогенолиз и глюконеогенез) на уровне, покрывающем потребности тканей, облигатно зависимых от нее, особенно мозга; 2) увеличение мобилизации главных энергетических запасов (жир); 3) уменьшение расходования белков организма. Нормальную реакцию на голодание можно рассматривать как непрерывный процесс, который удается подразделить на три фазы: состояние натощак (6— 12 ч после приема пищи), кратковременное голодание (в течение 12—72 ч) и длительное голодание (2 нед и более) [14]. После ночного голодания (состояние натощак) печень усиленно продуцирует глюкозу, которая потребляется в основном мозгом. Скорость кругооборота глюкозы у взрослого человека в этом состоянии составляет 2—3 мг/кг в минуту. Продукция глюкозы печенью определяется гликогенолизом (который обеспечивает 75% выхода глюкозы) и глюконеогенезом (за счет которого продуцируются остальные 25%). Основными субстратами, используемыми печенью для глюконеогенеза, служат продукты гликолиза — лактат и пируват, аминокислоты, особенно аланин, и глицерин. В состоянии натощак глюкоза в организме человека потребляется почти исключительно мозгом. Небольшие количества ее потребляются также форменными элементами крови. Совершенно иная ситуация характерна для инсулиночувствительных тканей, таких, как мышцы и жировая ткань, которые практически не потребляют глюкозу при голодании, длящемся 12 ч и более. Главным гормональным сигналом, обеспечивающим плавный переход от состояния сытости к состоянию натощак без развития гипогликемии служит снижение уровня инсулина в плазме. Если через 1—3 ч после еды уровень его повышается до 30— 100 мкЕД/мл, то в состоянии натощак он снижается до 10— 20 мкЕД/мл. Именно эта исходная (а не после еды) концентрация инсулина является условием стимуляции процессов гликогенолиза и глюконеогенеза, обеспечивающей такую продукцию глюкозы печенью, которая соответствует сохраняющимся потребностям мозга. Кроме того, уменьшение концентрации инсулина до исходной служит сигналом для прекращения потребления глюкозы инсулиночувствительными мышечной и жировой тканями. Если голодание продолжается более 12—14 ч, то включаются дополнительные механизмы сохранения эугликемии. Поскольку после ночного голодания запасы гликогена в печени составляют всего 70—90 г, а мозг утилизирует глюкозу со скоростью 125 г/сут, то при 24—48-часовом голодании печеночные запасы гликогена должны оказаться исчерпанными. В связи с этим поддержание печеночной продукции глюкозы и эугликемии требует прогрессивного нарастания глюконеогенеза в печени. Этот процесс в свою очередь зависит от мобилизации аланина и других аминокислот из мышечных запасов белка. Поглощение и превращение аланина в глюкозу в печени прогрессивно нарастает по мере увеличения длительности голодания более 12 ч. Кроме того, увеличивается и мобилизация жирных кислот из жировой ткани (т. е. стимулируется липолиз), что обеспечивает энергетическим материалом печень и мышцы. Помимо этого окисление жирных кислот в печени приводит к кетогенезу и прогрессивному повышению уровня b-оксибутирата и ацетоацетата в крови. Главным гормональным сигналом, обусловливающим увеличение мобилизации аминокислот, стимуляцию глюконеогенеза, липолиза и кетогенеза при длительности голодания более 12—14 ч, служит дальнейшее уменьшение концентрации инсулина в плазме ниже 10 мкЕД/мл. В генезе глюконеогенной реакции принимают участие также повышение содержания глюкагона в плазме и сохранение исходного уровня кортизола и гормона роста. Что касается механизма уменьшения концентрации инсулина в плазме, то главным стимулом для этого служит небольшое (на 150—200 мг/л) снижение содержания в ней глюкозы, которое обычно характеризует 24—72-часовое голодание. Однако отрицательный калорический баланс может и сам по себе обусловливать уменьшение секреции инсулина. Об этом свидетельствуют наблюдения, в которых инфузия небольших количеств глюкозы голодающим людям предотвращала снижение уровня сахара в крови, но не препятствовала развитию гипоинсулинемии [15]. Общие субстратно-гормональные нарушения, наблюдаемые при 14—72-часовом голодании, суммированы на рис. 11—2. Если голодание продолжается в течение нескольких дней или недель, то включаются гомеостатические механизмы не только поддержания эугликемии, но и сохранения белковых запасов орга-
Рис. 11—2. Гормональные и субстратные изменения, поддерживающие-эугликемию (и предотвращающие гипогликемию) у здоровых лиц при голодании. Главным гормональным сдвигом является снижение уровня инсулина в плазме, что приводит к усилению продукции глюкозы и снижению ее утилизации. Уменьшение содержания инсулина в плазме в свою очередь обусловливается небольшим снижением уровня глюкозы в ней (на 50— 100 мг/л) и/или самим по себе уменьшением поступления калорий. низма. Значение последних механизмов иллюстрируется тем фактом, что смерть при голодании является следствием расходования от 1/3 до 1/2 белковой массы тела, что обусловливает слабость дыхательной мускулатуры, ателектазы, пневмонию и сепсис. Поскольку белок служит единственным источником неуглеводных предшественников глюконеогенеза (печень млекопитающих не может превращать в глюкозу жирные кислоты), снижение распада белка неминуемо приводит к уменьшению интенсивности глюконеогенеза. Несмотря на снижение продукции глюкозы, эугликемия и удовлетворение потребностей мозга в энергетических субстратах сохраняются потому, что при очень длительном голодании мозг окисляет в качестве альтернативного субстрата кетоновые тела [16]. Важнейшей детерминантой утилизации кетоновых тел мозгом является концентрация этих соединений в артериальной крови. Точный уровень кетокислот и продолжительность голодания, при которых начинается утилизация кетоновых тел мозгом, неизвестны, но ата продолжительность наверное превышает 3 сут и, вероятно, меньше 14 сут. Дата добавления: 2015-02-05 | Просмотры: 1232 | Нарушение авторских прав |