|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
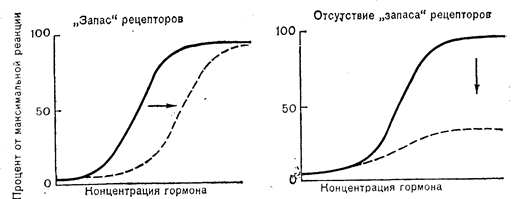
ВЛИЯНИЕ РЕГУЛЯЦИИ РЕЦЕПТОРОВ НА КЛЕТОЧНЫЕ РЕАКЦИИИз представлений о регуляции рецепторов следует, что изменение их концентрации должно влиять на реактивность клеток-мишеней. В отсутствие других изменений следовало бы ожидать, что уменьшение количества мембранных рецепторов приведет к снижению чувствительности клеток-мишеней в тканях, в которых есть большой «запас» рецепторов, и к снижению величины реакции в клетках-мишенях, в которых именно концентрация рецепторов становится лимитирующим фактором в отношении способности гормона вызывать реакцию (рис. 4—7). Общая функция регуляции рецепторов как механизма снижения чувствительности клеток-мишеней при повышении концентрации гормонов могла бы придавать организму определенные биологические преимущества. Наглядным примером является гомеостаз глюкозы, когда уменьшение числа инсулиновых рецепторов и снижение чувствительности периферических тканей к инсулину должно было бы «притуплять» влияние гиперинсулинемии на концентрацию глюкозы в крови. Регуляция рецепторов могла бы также сводить к минимуму или смягчать сердечно-сосудистые реакции на избыточные количества ангиотензина II и катехоламинов. Именно это могло бы определять выраженную резистентность к ангиотензину II, которая встречается при многих клинических состояниях, сопровождающихся повышенной секрецией ренина. Однако в отношении других стимуляторов, таких, как СТГ и гонадотропины, оказывающие на свои клетки-мишени более долговременные биологические влияния, задачи регуляции рецепторов не столь очевидны.
Рис. 4—7. Влияние потери рецепторов на биологические реакции (например, стероидогенез) в клетках-мишенях, обладающих и не обладающих «запасом» рецепторов.
Роль регуляции рецепторов в клеточной реакции на гормоны исследовали на различных тканях, особенно тех, в которых действие гормонов опосредуется системой аденилатциклаза — протеинкиназа. При изучении воздействия гормонов на такие ткани важно различать острую десенситизацию (обычно аденилатциклазы) без потери рецепторов и истинную потерю рецепторов, которая представляет собой отдаленное и медленнее обратимое следствие гормонрецепторного взаимодействия. Анализ уровня, на котором регуляция поверхностных рецепторов сказывается на метаболических реакциях, зависит от возможности измерения сдвигов по ходу активируемого гормоном процесса, «запускаемого» связыванием рецепторов. Для некоторых пептидных гормонов, таких, как инсулин, СТГ и пролактин,. ближайший механизм действия на клетки-мишени все еще не известен. Так, хотя инсулин модулирует активность нескольких мембранных ферментов, включая аденилатциклазу, фосфодиэстеразу и АТФазу, значение этих эффектов для действия гормона на мембранные транспортные системы и анаболические процессы остается не ясным. В том случае, если ранние этапы действия гормонов не идентифицированы, влияние регуляции рецепторов на клеточные реакции можно изучать путем регистрации сдвигов в транспортных системах и процессах биосинтеза. Такой подход применяли при анализе эффектов регуляции инсулиновых рецепторов в адипоцитах и гепатоцитах в отношении транспорта глюкозы и аминокислот, а также синтеза липидов. Последствия регуляции рецепторов в отношении функции клеток-мишеней широко изучали применительно к стероидогенным клеткам надпочечников и половых желез, реагирующим на тропные гормоны секрецией специфических стероидов, опосредуемой зависимым от аденилатциклазы механизмом. На таких клетках можно анализировать несколько уровней реакции, начиная с взаимодействия с рецепторами, продукции цАМФ и активации киназы и кончая стимуляцией продукции прегненолона и последующих этапов биосинтеза, приводящих к секреции стероидов. Если клетки получены после воздействия десенситизирующих доз тропного гормона, то in vitro регистрируются выраженные изменения содержания рецепторов и метаболических реакций на стимуляцию гормонами. Такие десенситизированные клетки демонстрируют ряд последствий регуляции рецепторов, осуществляемой действием гормона в интактных тканях-мишенях, в том числе снижение образования цАМФ и повреждение процесса биосинтеза стероидов [22].
Дата добавления: 2015-02-05 | Просмотры: 1008 | Нарушение авторских прав |