|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Монокулярная депривачия■------------ Левая сторона
Правая» - • Левая
сторона О63 глам сторона
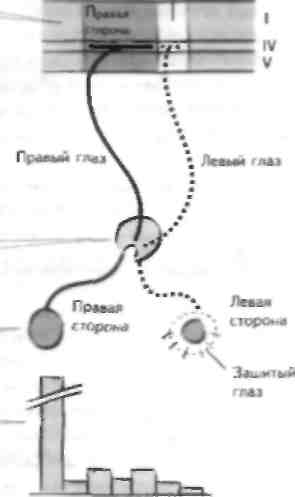
К настоящему времени получено достаточно дан-пых о том, что в ГДНС взрослого человека могут происходить регенерация и реорганизация. Однако пластичность в ГДНС опосредована не образованием новых нейронов, поскольку большинство нейронов в зрелой ГДНС находится в постмитотическом состоянии, а способностью к образованию новых аксонов. Наиболее выражена эта способность в раннем постнатальном периоде, когда все системы мозга развиваются, и именно тогда происходят наиболее значимые модификации. Механизмы, лежащие в основе пластичности, полностью не изучены, однако важную роль в этом играют продукция и усвоение факторов, способствующих росту нейронов и их выживанию (неиротрофические факторы). Пластичность развивающегося зрительного анализатора В первых исследованиях Хубель и Вайзель показали. что при рождении волокна поступают в IV слой первичной зрительной коры (ПЗК) диффузно и лишь в критический период развития (у кошек это составляет 3 14 нед, у человека несколько лет после рождения) зти волокна разделяются и распределяются по принципу селективности по стороне (см. главу 25). Разделение волокон зависит от типа импульса-ции по афферентным волокнам от каждого глаза; чем она больше, тем выше вероятность, что эти афферентные волокна будут регулировать определенную группу корковых нейронов. Так, селективность по стороне достигается в отсутствие какого-либо состя- зания между афферентными волокнами, идущими от глаз, однако она не может быть достигнута при отсутствии афферентного влияния хотя бы с одного глаза. Хубель и Вайзсль в эксперименте манипулировали этими волокнами, лишая один глаз афферентной импульсаиии путем сшивания его век (монокулярная депривация), а в последующих экспериментах проводили обратную процедуру (расшивание). Монокулярная депривация приводила к распространению таламического влияния от открытого глаза на IV слой, при этом впоследствии столбики клеток правого глаза реорганизовывались таким образом, что большинство клеток коры контролировалось открытым глазом. Это можно было изменить, расшив ранее закрытый глаз в критический период, что приводило к изменению таламического влияния на клетки коры, вызываемому активацией синапсов, которые ранее были функционально инактивирова-ны. Это было возможно, поскольку времени на окончательное прорастание аксонов к проекции было недостаточно. Однако в дальнейшем изначально инактивированные синапсы зашитого глаза будут утрачены, поскольку активирующие таламичес-кие волокна станут полностью контролировать корковые клетки. Правильное распределение импульсов в ПЗК и принцип селективности по стороне важны для развития многих других зрительных функций ПЗК. Однако по прошествии критического периода возможности реорганизации зрительной коры уменьшаются, по не исчезают. Пластичность у взрослых Анализатор общей чувствительности и окуловестибулярные рефлексы В настоящее время известно, что чувствительный анализатор может перестраиваться при нарушении афферентной импульсаиии от периферических рецепторов. Так, утрата имнульсации ОТ пальца (па-пример, при ею ампутации) не ПРИВОДИТ к инактивации зоны коры. Соседние отделы коры, получающие чувствительную импульсацию от других пальцев, распространяют свои аксоны на инактиви-рованпую зону коры. В противоположность этому избыточная афферентная чувствительная информация приводит к расширению корковой зоны, реагирующей па нее. Можно предположить, что активность одного афферентного волокна индуцирует продукцию нейротро-фических факторов в постсинантической клетке, которые затем связываются с соответствующими рецепторами на активной пресинантической терминал и, что приводит к росту и поддержанию ее функции. В этом смысле ЦНС постоянно перестраивается в зависимости от поступающей в нее афферентной информации. Впоследствии было показано, что выраженные чувствительные выпадения, например деафферента- пия конечное)и, приводи! к аналогичным изменениям, т.е. перестройке отделов коры за счет соседних импульсов, и это ЛОС1ШНС1СЯ не только за счез локального прорастания аксонов из соседних отелов коры. Друз им примером пластичности 1[П(являются окуловестибулярные рефлексы (см. главы 28 и 37). Вестибулярный анализатор подаез в ППС сигнал об ускорении движения головы, который затем проводится в мозжечок по моховидным волокнам. Однако волокна другого типа — ползущие — дают в мозжечок информацию о том, насколько при этом смешается изображение предметов на сетчатке (т.е. о том, насколько движения глаз могут компенсировать перемещение головы). Импульсация по ползущим волокнам нужна для того, чтобы проводить не только информацию о том, насколько хорошо реализуется данный рефлекс, но и импульсацию, с помощью которой рефлекс изменяется. Так, если изменить соотношение между информацией о положении глаз и головы, надев на человека призмы, рефлекс адаптируется к этим изменениям, это возможно благодаря тому, что импульсация по ползущим волокнам может модифицировать импульсацию по параллельным волокнам (и косвенно по моховидным волокнам) к клеткам Пуркннье посредством такого молекулярного механизма, как отсроченная депрессия (ОД; см. i iaej 37). Стволовые клетки Во множестве зрелых тканей гибель клеток в результате CC1CCIвенных процессов старения компенсируется последующей их пролиферацией и лиффе-ренцировкой. Считалось, что в ЦНС взрослого чс-иовека ною не происходит, однако недавно было показано, что в зрелой \\\\С млекопитающих имеются невральные клетки-предшественники. Эти клетки обнаружены в гиппокампс и паравентрику-лярно (в субвентрикулярной зоне), и, по-видимому, из них образуются функционально активные нейроны, однако их роль в пластичности и восстановлении до конца не изучена. Ограничения восстановительных возможностей ЦНС взрослого человека Необходимо представлять, что возможности восстановления \{ИС ограничены, потому что: • нейроны ЦНС находятся в постмитогическом состоянии, количество стволовых нервных клеток ограничено и они расположены только в определенных местах; • глиальные клетки ЦНС тормозят рост аксонов (см главу 4). Астроциты образуют сигналы, которые угнетают рост аксонов, а олигодендроциты продуцируют ряд факторов, приводящих к остановке роста аксонов. Это проявляется при поражении ЦНС, когда глиальные клетки начинают делиться и образуют рубец. ЧАСТЬ V. Заболевания нервной системы ГЛАВА 48. Неврологическое обследование
Дата добавления: 2015-02-06 | Просмотры: 963 | Нарушение авторских прав |

 Височная доля (речь, память. сложная зрительная информация)
Височная доля (речь, память. сложная зрительная информация)