|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Нарушения регуляции движений и локомоции спинного мозгаУ экспериментальных животных локомоция осуществляется без супраспинальных влияний (фиктивная локомоция), у людей такого не происходит. Однако нарушения ходьбы могут развиваться в силу различных причин. Нарушения функции ИН спинного мозга приводят к развитию синдрома ригидного человека (встречается редко). При этом заболевании у больных отмечается диффузная ригидность аксиальной мускулатуры со спазмами, вызванными им-пульсацией МН в результате отсутствия тормозных влияний на вентромедиальные МН, или без них. Синдром ригидного человека связан с наличием аутоантител к ферменту, синтезирующему гамма-аминомасляную кислоту (ГАМК), глутаматдекар-боксилазу (ГДК). Поражение нисходящих проводящих двигательных путей также наблюдается при некоторых заболеваниях. Наиболее тяжелое состояние отмечается при распространенном поражении ствола мозга, когда у больных утрачивается тормозное влияние на разгнбатсльную мускулатуру и формирустСЯДецереб-рационная ригидность с дугообразным выгибанием шеи и повышением мышечного тонуса в разгибателях конечностей. В противоположность этому более рострально расположенный очаг в одном из полушарий приводит к развитию контралатсралыюго гс-минареза млн к-мпнлеши с повышением мышечного тонуса и повышением сухожильных рефлексов О ннсррсфлскспя) (. ненропзво иными |чп мичпыми сокращениями мышц, спонтанными или в отпет на растяжение (клонус), что также Наблюдается при поражении нисходящих двигательных путей сипи ного мозга (см. главы 31, 32 и 41). При страдании пнрампдною факта ранпбателп поражаются и большей степени, чем ешбателн в руке, а в ноге возникает обратная ситуация. Это напевается пирамидным распределением парезов, что не совсем корректно, поскольку поражение пирамидных путей у обезьян приводит лишь к нарушению движений пальцев с некоторой степенью мышечной ыпнмоппп и пнюреф.текепп. ГЛАВА 35. Двигательные зоны коры
I Медиальная часть полушария мозга I Дополнительная моторная зона Первичная моторная кора
Поясная борозда Префроиталь-ная кора Двигательные корковые отделы I Периферический/соматосенсорный афферентный импульс — Зрительные отделы коры л Передние отделы Префрон--------
поясной извилины тальная кора I___ I
В регуляции движений участвует несколько отделов коры: первичная моторная кора (см. главу 36), премоторная кора (I I I'M К), дополнительная моторная ЗОНа (ДМЗ), а также ряд соседних областей в передних отделах поясной пошлины. Кроме того, существуют отделы мозга, участвующие в корковом контроле движений, например корковый центр ВЗОра (см. главу 40) и чадняя теменная кора (см. главу 30). В Этой главе кратко обсуждаются организация двигательных отделов коры и их роль в регуляции движений, в следующей главе речь пойдет преимущественно о первичной моторной коре. Первичная моторная кора Первичная моторная кора (ПМК) - это часть коры мозга, которая реагирует двигательным ответом на минимальную электрическую стимуляцию. Она соответствует полю 4 по Бродману, располагается кпе- рели о'| центральной Оорозды и проецируется на МП ствола мозга посредством кортиконуклелрных путей и ни МП спинного МОЗГа посредством кортикоспи- 11.1.a,in,i\ трактов (КС Т), а также непрямым способом на подкорковые экстрапнрамндные тракты. Действительно, ПМК особенно тесно связана с пирамидным трактом (хотя 60 70/г его начинается от других отделов корм) и поэтом) принимает участие в регуляции тонких движений дистальной мускулатуры (главы 34 п 36). Другие корковые отделы В регуляции движений принимает участие несколько отделов коры: ИРМ К (латеральная часть поля 6 по Бродману), ДМЗ (медиальная часть поля 6 но Бродману), некоторые корковые отделы передней части поясной извилины, медиальная часть лобной доли, корковый центр взора (поле 8 по Бродману) и задняя теменная кора (поле 7 по Бродману). Некоторые ИЗ этих отделов имеют особые функции. Например, корковый центр взора отвечает за движения глаз (см. главу 40), а задняя теменная кора осуществляет зрительный контроль за движениями (см. главу 30). Остальные отделы лобной доли регулируют более сложные движения. Большинство этих корковых отделов занимают большую часть двигательной иерархии, нежели ИМ К, их связи и функции представлены в табл. 35.1 (см. главу 32). И ГМК относится к полю 6 по Бродману и. как и первичная моторная кора, посылает импульсы непосредственно к МН по КСТ или пирамидному тракту. Этот отдел занимает два уровня двигательной иерархии, поскольку участвует в планировании двигательных актов (см. главу 32). В противоположность этому ДМ 3 принимает участие в планирова- нии движении, особенно г. on.ii на сенсорные СТИМУЛЫ. Более того, в настоящее время стало известно, чтоДМЗ — л ишь часть большого количества двигательных корковых отделов, лежащих на медиальной стороне лобной доли и вовлеченных в большей степени в процесс планирования, нежели выполнения движений. Именно эти отделы коры получают импульсацию от базальных ганглиев (см. главу 38), что позволяем обьясипч ь аномальные движения, наблюдающиеся при заболеваниях этого отдела мозга человека (см. главу 39). Например, при болезни Пар-кинсона отмечаются замедление и обеднение двигательного рисунка, которые связаны с недостаточной активностью этих корковых отделов и устраняются с помощью противопаркннсоническнх препаратов или нейрохирургического вмешательства.
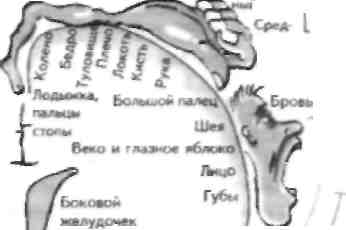
ГЛАВА 36. Первичная моторная кора
Дополнительная моторная зона (ДМЗ) Первичная Медиальная часть полушария головного мозга ДМЗ
теменная кора Поясная ЗВИЛИКЭ Связи двигательной коры Импульсация от других двигательных зон
Премотор-ная кора Язык/А / *V поганее//*. Саливация Латеральные отдели
Жевание
^Кожные рецепторы 'Суставные ^рецепторы ^Мышечное веретено Мышцы, сгибающие палец МН спинного мозга, иинераирующий сгибатели пальца
ли кор-, тракт I 5% крупнокалиберных воло | кон КСТ исходят из пирамидных клеток Беца и V слоя ПМК
Первичная моторная кора (ПМК) получаст афферентную информацию ОТ мозжечка (через тала-мус), от Отделов коры, расположенных кпереди от нее, а именно дополнительной моторной зоны (ДМЗ) чувствительные импульсы от мышечных веретен и сенсорных областей коры. Последнее обстоятельство указывает на то, что разделение отделов ЦНС на двигательные и чувствительные является условным. Поэтому для удобства понимания необходимо выделять ПМК и первичную соматосенсор-ную кору (ПССК) (см. главу 20). Изучение ПМК показало, что двигательная иннервация тела человека организована соматотонически, КО при ном корковое предстаг.шельство движений и каждом отделе тела человека пропорционально степени двшачелыюп иннервации, например мускулу-тура руки и лица имеет наибольшее представительство. Обратная соматотоннческая проекция метафорически называется двигательным гомункулом, у которого голова представлена в латеральных отделах коры, а ноги — в медиальных отделах. В соответствии с этой соматотопической проекцией у больных возникают эпилептические припадки при раздражении моторной коры, при котором они могут начаться на одной стороне, обычно в руке, а затем распространиться в виде клонических подергиваний на другие отделы (джексоновский марш, названный по имени описавшего его невролога Х.Джексона). В противоположность этому припадки, возникающие при раздражении ДМЗ, начинаются с поднимания рук и вокализапии, что указывает на большую значимость этой области в регуляции движений (см. главу 35). Исследования соматотопической проекции Пен-филдом и соавт. в 50-х годах прошлого века позволили установить макроскопическую организацию ПМК, но впоследствии исследования с микроэлектродами у животных показали, что ПМК организована в виде столбиков двигательных нейронов (см. главу 15). Импульсация к этим столбикам поступает по афферентным волокнам от суставов, мышечных веретен и кожи, и они максимально активируются при сокращении мышц, активируемых этим же отделом коры. Так, например, группа столбиков двигательных нейронов получает сенсорные стимулы от пальца, когда он согнут, эта импульсация проводится рецепторами кожи па передней поверхности п&ть-ца — мышечными веретенами мышц, сгибающих палец, а также суставными рецепторами суставов пальцев. Таг же самый столбик двигательных нейронов посылает импульсацию к МП спинного мозга, которые ипперпнруют сгибатели пальца. Активация кортикоспиналыюго нейрона в этом столбике активирует рецепторы, которые проецируются на этот же столбик, и наоборот. Считается, что каждый столбик получает прямую и обратную афферента-цню, что имеет важное значение для рефлекторного контроля за движениями, допустим, па примере длиннолатентных, или транскортикальных, рефлексов. 'Оти рефлексы приводят к отсроченным и мало-выраженным элсктромнографическим (^МГ) изменениям, которые возникают после растяжения мышцы и первым ЮМГ-измепсннсм будс'1 М ответ при вызывании мопосипаитическорго рефлекса на растяжение (см. главу 33). Афферентным компонентом траНСКОртиКаЛЬНОГО рефлекса является афферентное волокно от мышечного веретена 1а (импульсация передается по задним столбам и медиальной петле), а эфферентным его компонентом — корти-коспинальный тракт (КСТ). Роль этого рефлекса неизвестна, но он имеет важное значение в корковом контроле движений, сообенно если в процессе двигательного акта человек наталкивается на препятствие, которое активирует мышечные веретена. Имеются противоречивые данные о том, осуществляет ли П МК непосредственный контроль за конкретными мышцами, простыми движениями или же некоторыми компонентами движений. Нейроны ПМК реагируют еще до появления ЭМГ-изменений и. вероятно, колируют направление и силу движения, несмотря на то что эта активность зависит от харак-iepa выполняемою действия. Поэтомув целом двигательная кора контролирует движения, иннервируя определенные МП (конкретные кортикоспинальные аксоны иннервирукп множество различных МП). ПМК может ремоделнроваться после очаговых поражений или изменений сенсорных афферетных ешмулов, что говорн'1 о ее способное!и поддерживать гибкое сообщение с мышцами в течение жизни. Гак, нейроны ПМК могут начать регулировать другие мышцы. В опредленных отделах коры возможно изменение еннашнческой активности с помощью отсроченной позенцнацн» (см. главу 45), это свидетельствует о том, что ПМК может принимать участие в обучении новым движениям, т. е. обладать функцией, традиционно приписываемую мозжечку (см. главу 37). Изолированное поражение ПМК встречается редко и в эксперименте, как правило, приводит к нарушениям, сходным с таковыми при страдании отдельных частей пирамидного тракт. Однако поражение ПМК с обеих сторон в сочетании с поражением прсмоторных зон, что наблюдается при ише-мическом инсульте в бассейне средней мозговой артерии (см. главу 17), приводит к более выраженному неврологическому дефициту, в частности развитию гемниареза. В противоположность этому, поражение ДМ'З приводит кдвусторонним нарушениям инициации движений, toi да как поражение коркового центра взора к ограничению взора. ГЛАВА 37. Мозжечок Макроанатомия мозжечка Г^ яем*1мческии уплощенный вид моаже^ка сзади]
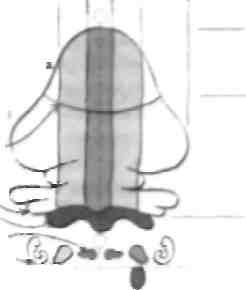
4. Импуяясацив по ползущим волокнам исходит из I S- Гломерулы иоэжечк» содержат моховидные волокна, котором* лают ым-пугмсацнто ГК 6. Глубинные «два мозжечка тонмчас к* активируются коллатеральными моаовидиымн волокнами н попзутцм-ми волокнами, что поззоляат осуьнь ствлягь c 7 Импу/мСацмя ОТ КЛ К глубинным ядрам мозжечка носит тормозной характер
Анатомия мозжечка Мозжечок ч<> сложная структура, которая ми ходится под наметом твердой мозговой оболочки в задней черепной ямке и связана со стволом мозга тремя нарами ножек мозжечка (см. главу 13). Мозжечок отвечает за координации) движений и обучение двигательным навыкам. В мозжечке имеются 3 функциональные и анатомические системы: • спипоцеребеллярпая участвуем и регуляции функций аксиальной мускулатуры и поддержании позы; • понтоцеребелляриая участвует в координации и планировании движений конечностей; • вестибулоцсрсбслляриая участвует в поддержании позы и движении глаз. Эти 3 системы имеют собственные связи (см. Приложение 3). Спиноцеребеллярная система подразделяется на вермальный и паравермальный (промежуточный) отделы, при этом последний тесно связан с аксиальной мускулатурой, а также с вентромедиаль-ными нисходящими двигательными проводящими путями и мотонейронами, тогда как паравермальная часть в большей степени отвечает за координацию дви- жений. Понмоцеребсллярная система участвует в координации движений, но в большей степени связана со зрительным контролем за движениями и передает информацию ом задней меменной коры к двигательным о/делам коры. Несмибулоцерсбсллярная система не ассоциирована е глубокими ядрами мозжечка и является одной из филогенезически наиболее древних частей мозжечка. Как и перма.п.ная часть спиноцсре беллярной системы, она связана с венмромелиа.мьны-ми двигательными путями, но при этом участвует в ре1уляции движений глаз. 15 общем, мозжечок сравниваем движение, комо-рое человек намереваемся сделать, информация о котором поступаем из двигательных областей коры, с денем вимсльпым движением, информацию о КОТОРОМ передаю! афферспмные волокна мышечных веремеп и интернейроны спинного мозга. При этом мозжечок получает информацию от вестибулярного и зрительного анализаторов. Когда сравнение проведено, сигнал о погрешности поступает по нисходящим проводящим двигательным путям и корригирующий фактор сохраняется в виде двигательной памяти, опосредованной синоптической имнульсанией к клеткам Пуркиньс (КП). Эти модифицируемые синапсы на уровне КП являются примером отсроченной депрессии (ОД; см. главы 45 и 47). ОД приводи'! к уменьшению синаптнческой импульсации параллельных волокон (ИЛИ) к КП, когда импульсация активирована по фазе и низкой частоте ползущим волокном (НОВ). Этот процесс происходит в течение как минимум нескольких часов. Другими словами, в период совершения новых движений импульсация к КП по ползущим волокнам увеличивается, что оказывает модулирующее влияние на импульса -цию по параллельным волокнам к той же КП. Когда движение усваивается, импульсация по ползущему волокну уменьшается, но возрастает модулирующее влияние по параллельному волокну; считается, что именно этот механизм лежит в основе обучения двигательным навыкам. Гипотеза подобной модулируемой синаптнческой импульсации была впервые предложена Марром в 1969 г. и впоследствии подтверждена, сообенно при изучении окуловестибулярных рефлексов (см. главы 28 и 47). Биохимические механизмы ОД в мозжечке не полностью изучены, но, по-видимому, в их основе лежит активация различных глутаматных рецепторов КП и последующий приток кальция и активация протеинкиназ. Модулируемая синаптическая импульсация отвечает за обучение и хранение информации, относящейся к двигательной памяти (см. Приложение 3). Микроскопическая структура мозжечка, обеспечивающая генерацию ОД, хорошо изучена, несмотря на то что биохимические механизмы изучены недостаточно. Возбуждающие импульсы в мозжечок проводятся моховидными и ползущими волокнами. Эти моховидные волокна непрямым образом активируют КП через ПАВ, которые исходят из гранулярных клеток (I К). В Противоположность этому НОВ образуют прямые синапсы с КП и, как в случае импульсации по моховидным волокнам, посылают импульсы к нейронам глубинных ядер мозжечка (111 ЯМ). Эти нейроны тонически возбуждаются афферентными волокнами, проводящими импульсы в мозжечок, а зачем ингибируются корой мозжечка, т. е. КП. В свою очередь КП ИНГИби-руются некоторыми локальными иптернейронами, тогда как клетки Гольджи (КГ) в наружном гранулярном слое посылают тормозные импульсы к ГК. Эти интер-пейропы способны тормозить субмаксимально активированные КП и ГК и таким образом проводить именно тот сигнал, который необходимо проанализировать. Итоговая импульсация от глубинных ядер мозжечка на структуры ствола также является тормозной. Дата добавления: 2015-02-06 | Просмотры: 1539 | Нарушение авторских прав |



 Центральная борозда
Центральная борозда , Дополнительная (моторная зона (ДМЗ)|
, Дополнительная (моторная зона (ДМЗ)|

 Полушарие головного мозга (вид сбоку)
Полушарие головного мозга (вид сбоку) ПМК Задняя
ПМК Задняя Сенсорные стимулы
Сенсорные стимулы

 Параллельное волокно
Параллельное волокно