|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Получает мелкокалиберные немиелинизированные волокна от комплекса верхних оливСлуховой анализатор отвечает за восприятие звуков. Главным рецептивным органом является улитка во внутреннем ухе, которая преобразует звуковые волны в электрические импульсы с помощью процесса механического проведения. Электрический импульс, генерируемый в ответ на звук, проходит (вместе с информацией от вестибулярной системы; см. главу 28) через черепно-мозговой нерв (ЧМН; преддвер-но-улитковый нерв) в ствол мозга, где образует синапсы с комплексами кохлеарных ядер (см. главу 27). Несмотря на то что слуховой анализтор выполняет много функций, первичный анализ звукораз-личения происходит в улитке. Свойства звуковых волн Звуковая волна характеризуется амплитудой и громкостью (измеряемой в децибелах дБ), частотой (измеряемой в герцах Гц), формой, фазой и качеством илитембром. Интенсивн ость звука варьирует очень широко, но обычно различается звук интенсивностью 1—2 дБ. Поступление звуковой волны создает разность фаз и интенсивности между обоими ушами, если только звук не исходит из средней линии. Степень задержки и изменения интенсивности между обоими ушами в результате физического разделения необходима для определения локализации источника звуков (см. главу 27). Наружное и среднее ухо Достигая уха, звук проходит по наружному слуховому проходу к барабанной перепонке, которая вибрирует с частотой и силой, определяемыми звуком. Это приводит к смещению трех слуховых косточек в среднем ухе, смещению жидкости в улитке при внедрении отростка стремечка в овальное окно улитки. Этот процесс необходим для уменьшения звукового сопротивления и усиления реакции на звук, поскольку звук воздействует на жидкость непосредственно с сильным отражением. СлуХОВЫМН косточками управляют две маленькие мышцы, которые предохраняют косточки от повреждения ГРОМКИМИ звуками, а также изменяют положение отростка стремечка в овальном окне. Повреждение слуховых косточек (отосклероз), среднего уха (среднийотит) или наружногослуховогопрохода (закупорка серной пробкой) приводи! к снижению остроты слуха или тугоухости, которая по своей природе является кондуктивной. Внутреннее ухо и улитка Смещение ОТрОСТКа стремечка в овальном окне приводит к генерации волны в заполненной пе-рилимфой лестнице преддверия и барабанной полости улитки. Эти две лестницы сообщаются с апикальным краем улитки — геликотремой, но на всем остальном протяжении разделены срединной лестницей, которая содержит звукопроводящий аппарат, или кортиев орган. Кортиев oprt.......... аходизся на дне леезницы на структуре, называемой базальной мембраной (ЬМ), ширина козорой увеличивается с увеличением рас стояния от конца стремечка. Увеличение ширины связано со снижением плотности БМ, это необходимо для того, чтобы звуки высокой частоты максимально смещали БМ на конце стремечка улитки, а звуки низкой частоты максимально активировали апикальный конец БМ. Так, различение частот является одной из функций БМ. несмотря на то что эта способность значительно увеличивается при помощи волосковых клеток кортиева органа, который лежит на этой мембране. Кортиев орган является сложной структурой, состоящей из клеток слухового восприятия - волосковых клеток (см. главу 19), которые по своей структуре подразделяются на два типа: однорядные внутренние волосковые клетки (ОВВК), дающие большую часть импульсации в преддверно-улитко-вый нерв, и 3—4-рядные наружные волосковые клетки (НВК), регулирующие реакцию ОВВК на звук. Эти два типа волосковых клеток отличаются морфологически и электрофизиологически. ОВВК получают небольшую проекцию от ствола мозга, тогда как ПИК получают импульсапию ОТ комплекса верхних олив, который влияет на форму и способность клеток воспринимай, звуки. Некоторые ОВВК непосредственно контактируют с текториачыюй мембраной (ТМ) кортиева органа, что важно для регуляции реакции НВК на звук, так как эти клетки не контактируют с'Г М, но обеспечиваю! 93S* импульсации по преддверно-улитковому нерву. Одно афферентное волокно получает импульсапию ОТ множества ОВВК, однако лишь одна II ВК связана с несколькими афферентными волокнами. Кроме того, есть и другие различия между ОВВК и НВК. Они неоднородны на протяжении срединной лестницы. Эза неоднородность клеток по форме изменяет их частотно-воспринимающие характеристики, что позволяет клеткам до определенной степени настраиваться на частоту звука, помимо опосредования этого процесса свойствами БМ. Повреждение улитки, волосковых клеток и кох-леарной порции преддверно-улнзкового нерва приводит к тугоухости, которая по природе является ней-росенсорной. Травма, ишемия и опухоли VIII ЧМП также приводят к нейросенсорной тугоухости. Недавно покачано, чзо наследезвенная глухоза связана с дефектами определенных белков стсреоцилий волосковых клеток. ГЛАВА 27. Слуховой анализатор II: слуховые пути и речь
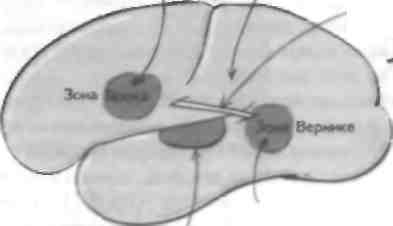
Речь Поражение аркуатных волокон приводит к акустико-мнестической афазии Аркуатиые волокна -^ Поражение зоны Вернике приводит к сенсорной афазии / • Первичная слуховая кора — поле 41 по Бродману • Вторичная слуховая кора — поля 41 и 42 по бродману
.Тонотопическая ' организация
'Столбики распределены в зависимости от связи с ушами через мозолистое тело Медиальное коленчатое тело т
>Т> Верхние бугор-1 1-1----- -в» ки четверохол-1
J — __ I
Комлекс верхних олив (КВО), возможно, ходи рует распознавание временных различий попадания звука в слуховой аппарат и локализации источника звука. Каждый нейрон КВО максимально активируется импулвсацией от каждого уха, прибывающей одномоментно на его аксо-иалъный бугорок, — компонент Л1 кодируется положением синапса, образованного волокнами от каждой улитки в зависимости от части нейрона, где он расположен. Кроме того, кодирование происходит в зависимости от длины аксонов от каждого кохлеарного ядра Голова создает уменьшение At (т. е. временную разницу) в достижении звуком каждого уха и снижение интенсивности д|. поскольку голова поглощает некоторое количество звуковой энерг ни. Мочка уха определяет локализацию источника звука в вертикальной плоскости. Предлверно-улитковый, или VIII, черепно-мозговой нерв перелает информацию от обеих улиток и вестибулярного аппарата (дальнейшее обсуждение см. главу 28). Каждое волокно улиткового нерва селективно настроено на определенную частоту, которая определяется по источнику звука внутри улитки (см. главу 26). Эти волокна затем организуются в зависимости от локализации иннервируемых ими во-лосковых клеток по длиннику базальной мембраны, и эта тонотопическая организация поддерживается в слуховых проводниках. На входе в ствол улитковый нерв образует синапсы с кохлеарными ядрами продолговатого мозга. Слуховые пути Кохлеарные ядра подразделяются на вентральное (ВКЯ) и дорсальное (ДКЯ). ВКЯ проецируется на комплекс верхних олив (КВО) с двух сторон, а ДКЯ через задние слуховые волокна на противоположное ядро латеральной петли и нижних бугорков четверохолмия. КВО содержит веретенообразные нейроны с латеральными и медиальными дендритами, которые получают импульсацию от каждого уха. Это первый уровень бинауральных взаимодействий, и эта структура играет важную роль в локализации источника звука. Медиальная часть КВО принимает возбуждающую импульсацию (ВВ-клетки). тогда как латеральная часть КВО принимает тормозную импульсацию от одного уха и возбуждающую — от другого (ВТ-клетки). ВВ-клетки принимают участие в локализации источника звуков низкой частоты (менее 1,4 кГц), при этом критерием является снижение частоты звука (М), поступающего в одно, а затем в другое ухо. На рисунке показано, как эти клетки проводят импульсы, генерируемые звуковыми сигналами различной локализации, к одному нейрону КВО от обоих ушей. ВТ-клетки играют важную роль в локализации источника звуков высокой частоты, причем критерием в данном случае является изменение интенсивности (AI) между ушами (при создании Д1 голова выполняет функцию щита). Анализ звуков с частотой выше 1,4 кГц (у людей) проводится по I. Гели источник звука находится па средней линии, то ни Д|, ни Д| не образуется и происходит как бы слияние источника локализации, которое до некоторой степени можно преодолеть, поменяв положение головы или подключив другие сенсорные стимулы. Локализация источника звука в вертикальной плоскости зависит от мочки уха. КВО не только проецируется рострально на нижние бугорки четверохолмия (II 1>Ч), по и носьиаст импульсацию к улитке, где контролирует ОВВК и изменяет воспринимающие свойства кортиева органа (см. главу 26). Проекция к IIБЧ является тоното-пической, и эта структура также получает импульсацию от первичной слуховой коры (ИСК) и других сенсорных модальностей. Она взаимодействует с верхними бугорками четверохолмия и участвует в пространственной ориентации в соответствии с по лучаемыми слуховыми стимулами (см. главы 24 и 40). НБЧ проецируются на медиальное коленчатое тело (МКТ), которое проецируется на ПСК в верхней височной извилине. Эти участки коры соответствуют полям 41 и 42 по Бродману, при этом тала-мическая импульсапия поступает преимущественно в III и IV слои коры. Столбиковая организация ПСК плохо изучена, но в соответствии с тонотопи-ческой проекцией низкочастотные звуки локализуются в задних отделах, а высокочастотные — в передних отделах. Вторичная слуховая кора и речь В ПСК, кроме нейронов, получающих относительно простые афферентные импульсы, находятся клетки, которые реагируют на некоторые сложные звуки. Эти нейроны в большей степени представлены во вторичной слуховой коре и наиболее сложно уст роен i,i у человека в зоне Вернике корковом центре узнавания речи. Эта область находится в доминантном полушарии (обычно левом), и при ее поражении (например, при инсульте; см. главу 17) возникает сенсорная афазия (логорея), или невозможность воспринимать обращенною речь. '')та область связана аркуатными волокнами с зоной Брока в лобной доле доминантного полушария, которая отвечает за экспрессивную речь; ее поражение приводит к развитию моторной эфферентной афазии, т. е. к отсутствию спонтанной речи при сохранности арти-куляторного аппарата. Тотальная афазия это полное отсутствие речевой продукции, которое наблюдается при инсульте в бассейне средней мозговой артерии с поражением зон Вернике и Брока. Селективное поражение аркуатных волокон приводит к развитию акустико-мнестической афазии, когда больные Moiyi понимать обращенную речь и творить, но не могут повторять слова и предложения. Однако в настоящее время получены данные о более сложных взаимодействиях между отделами мозга в процессе узнавании и речевой продукции. Афазию следует отличать от нарушений инициации и выполнения речедвшателыюго акта (например, паралич мягкого нёба и языка при болезни двигательного нейрона), которое называется дизартрией (или анартрией, когда речевая продукция полностью отсутствует). ГЛАВА 28. Вестибулярный анализатор
Ампулярный и«рв —"~^| | (Ампуяяриый гребень Дата добавления: 2015-02-06 | Просмотры: 1144 | Нарушение авторских прав |
 Угол наклона
Угол наклона Волосковал клетка полукружного канала I
Волосковал клетка полукружного канала I Смещение купулы приводит к смещению реснички волосковой клетки, окруженной этим веществом
Смещение купулы приводит к смещению реснички волосковой клетки, окруженной этим веществом