|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Нарушение функций ионных каналов в клинике нервных болезнейНекоторые лекарства оказывают фармакологическое действие на уровне ионных каналов. К ним относятся, например, местные анестетики и некоторые иротивоэпилептическис препараты. Однако в последние голы были описаны некоторые неврологические заболевания, протекающие с преимущественным поражением мыши и вызванные мутациями генов, кодирующих натриевые и хлорные ионные каналы. К ним относятся некоторые формы миотонни (нарушение расслабления скелетной мус-кулазуры после произвольного сокращения, приводящее к невозможности свободно выполнять те или иные двигательные акзы) и различные формы периодического паралича, при которых у пациентов развиваются приступы преходящей парциальной или генерализованной мышечной слабости. Волсс того, некоторые формы семейной гемиплегической мигрени или мозжечковых атаксий (см. главу 37) связаны с нарушением функции кальциевых каналов, а некоторые формы эпилепсии (см. главу 53) могут бызь вызваны нарушением функции специфических ионных каналов. Другие заболевания могут быть вызваны перераспределением или функпионирова пнем тех ионных каналов, которые в норме неактивны. Такие нарушения происходят в области, близкой к перехвачу Ранвьс, в результате центральной де-м иел ин и заци и при рассеянном склерозе или перифсричсскойдемислиннзации (синдром Пшена— Барре), что приводи! к нарушению распространения потенциала действия (см. главы 6 и 54). ГЛАВА 6. Мембрана в состоянии покоя и потенциал действия
активации потенциала
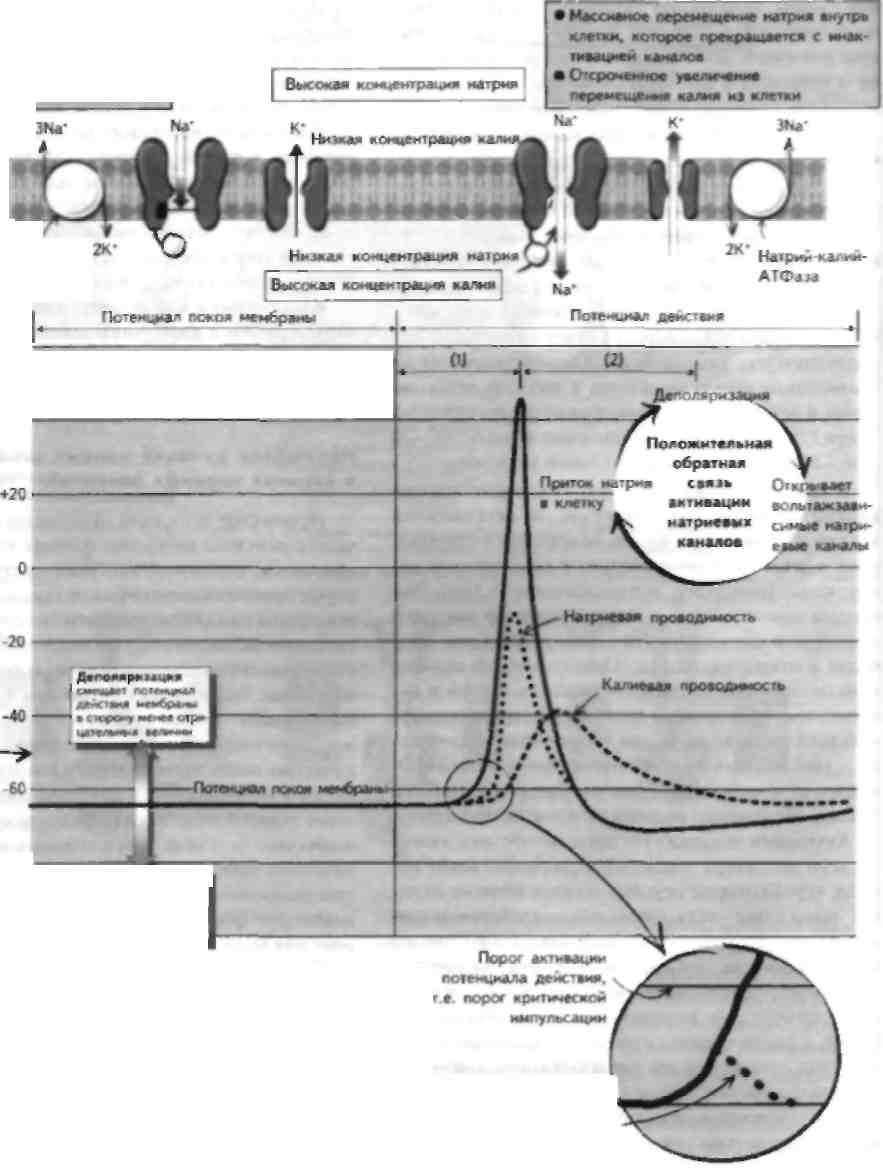
действия Так, например. £,* в скелетной мышце человека будет равен: Е.' -58 1л 3/140 - -95 мВ Потенциал покоя мембраны В покое мембрана нейрона относительно непроницаема для ионов. Это важно для генерации потенциала покоя. Основным интрацеллюлярным ионом является калий, а экстрацеллюлярным — натрий. Поэтому в соответствии с электрохимическим градиентом поток ионов калия направлен из кички, а поток ионов натрия в клетку. Перемещение положительно заряженных ионов из клетки приводит к генерации отрицательного мембранного потенциала или гиперполярнзации, тогда как поток ионов натрия внутрь клетки вызывает деполяриза- iini.i. Однако покоящаяся мембрана относительно непроницаема для натрия, но в то же нремя достаточно проницаема для калия. Поэтому и покос калий имеет тенденцию к перемещению из клетки в соответствии с электрохимическим градиентом, что приводит к отрицательному заряду мембраны. Гак продолжается, пока химический градиент концентрации, приводящий к выходу калия из клетки, не выключится разностью электрического потенциала, вызванного притоком ионов в клетку (мембранным потенциалом) и приводящего к возвращению калия в нейроны. Мембранный потенциал, обеспечивающий это стационарное состояние, называется потенциалом равновесия для калия (Е*) и вычисляется по уравнению Нсрнста (см. рисунок). На самом деле измеренный потенциал покоя аксонов имеет более положительную величину, поскольку незначительная проницаемость мембраны для ионов натрия, в покое сохраняется. Небольшой приток натрия поддерживается аденозинтрифосфат ЛТФ)-зависимой натрий-калий-АТФазой, которая сама по себе обладает небольшой электрогеннос-тью. Этот насос важен для поддержания ионных градиентов и является электрогенным, в силу того что обменивает 3 иона натрия на 2 иона калия, входящих в клетку. Это вносит лишь незначительный вклад в потенциал покоя мембраны. Генерация потенциала действия Одним ИЗ фундаментальных свойств нервной системы являются генерация и проведение нервных импульсов (см. главы 8 и 19). Эти импульсы включают генераторные, синаптичсскис потенциалы и потенциалы действия, последние определяют как одиночные электрические импульсы, распространяющиеся по аксону. Этот потенциал действия (нервный имиу.п.с или спайк) возникает по принципу «все или анчего», т. е. когда стимул достигнет определенного i трога интенсивности. Таким образом, информация II нервной системе закодирована по частоте имиуль-сации, а не по размеру потенциала действия (см. главу 18). Порог интенсивности стимула определяется как величина, до которой поднимается внутренний сетевой ток (зависимый в основном ОТ ионов натрия), которая выше, чем внешний сетевой ток (зависимый в основном от ионов калия), и которая составляет порядка 55 мН (критический порог им-нульеации). Он возникает в области аксонального бугорка, где плотность натриевых каналов наиболее высока и где инициируется потенциал действия нейрона. Однако если порог не будет достигнут, деполяризация не генерирует потенциал действия и сигнал не распространится по аксону. Последовательность событий, участвующих в генерации потенциала действия, такова: • Деполяризующий заряд активирует вольтажзави-симые натриевые каналы на мембране нейрона, Kulopi.ni Позволяет IICkulupUM UullaM HaipilH IK рсмсститься в соответствии со своим электрохимическим градиентом (повышенная натриевая проводимость). Это деполяризует мембран) И «и крывает сше больше натриевых каналов согласно механизму положительной обратной связи. Когда открывается достаточное количество натриевых каналов, для тою чтобы величина внутреннего тока сети превысила величину тока, обеспеченную выходом калия из клетки, открываются быстрые натриевые каналы, что приводит к массивному притоку натрия, который деполяризует мембранудо потенциала равновесия для натрия (примерно +55 мВ). Таким образом, генерируется спайк потенциала действия, однако он не достигает потенциала равновесия для натрия в силу продолжающегося и увеличивающегося оттока ионов калия из клетки. • Далее следует убывающая фаза потенциала действия, при этом вольтажзависимые натриевые каналы инактивируются (см. главу 5). Эта инактивация также вольтажзависима, однако она более медленная, чем фаза активации, и поэтому возникает позже (см. главу 5). Во время фазы убывания потенциала приобретает более важную роль вольтаж-зависимый калиевый поток, так как его активация деполяризацией мембраны происходит еще более медленно, чем инактивация натриевых каналов. Активированные вольтажзависимые калиевые каналы обеспечивают короткий период гиперполяризации мембраны, предшествующей ее дезактивации. и потенциал мембраны возвращается в состояние покоя. Сразу после появления спайка потенциала действия отмечается рефрактерный период, когда нейрон становится невозбудимым (абсолютный рефрактерный период) или же акт ивируется только субмаксимальными стимулами (относительный рефрактерный период). Абсолютный рефрактерный период возникает в момент максимальной инактивации натриевых каналов, тогда как относительный в более поздний период, когда большинство натриевых каналов уже вернулось в состояние покоя, но ток через активированные калиевые каналы уже раходится в развернутой фазе. Рефрактерный период содержит два важных аспекта для генерации и проведения потенциала действия. Во-первых, потенциалы действия могут распространяться только в одном направлении — от мест генерации, и, во-вторых, они могут генерироваться лишь с определенной частотой. Оригинальное описание потенциала действия принадлежит Ходжкину и Хаксли, которые изучали этот феномен на гигантском аксоне кальмара в 50-х годах XX века. Этот механизм был подтвержден позже и для многих других клеток и нейронов. Это наряду с открытием большого количества ионных каналов послужило началом описания многих других модификаций генерации потенциала действия нейронами и другими клетками. ГЛАВА 7. Нервно-мышечное соединение и синапсы
2 4099 16482 2151 17086 2151 17086 1947 17086 1947 17086 872 17086 872 17086 0 12452 0"> Дата добавления: 2015-02-06 | Просмотры: 1553 | Нарушение авторских прав |
 Порог
Порог олевание
олевание