|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Тело нейрона (сома) — это часть нейрона, которая содержит ядро и окружающую цитоплазму. В теле
нейрона происхолит обмен пенкстн и содержится большинство органелл нейрона (митохондрии, аппарат I о.п.лжн и пероксисомы). Кроме Iого, от. [ела Нейрона от\о;шт,'ir.a lima oi рое i koi.: денлриты II аксон. Многие нейроны имекп также базофилыюе содержимое. Субстанция Ниссля, состоящая из гранулярного эндоплазматического ретикулума и рибосом, отвечает за синтез белков. Она находится в цитоплазме тела нейрона и дендритах и отсутствует в аксональном бугорке и самом аксоне по не вполне ясным причинам. Кроме того, в теле нейрона и аксоне содержатся нейрофиламенты, которые поддер- кивают структуру цитосклелета нейрона. Более того, важно упомянуть дне другие фибриллярные структуры нейрона: микротрубочки и мнкрофнламенты, которые необходимы для аксоплазматического тока (см. ниже) и роста аксона. Дендриты это отростки нейронов, которые отходят от тел нейронов, ветвятся и отвечают за передачу информации к телу нейрона от синапсов дендритического дерева (аксодендритические синапсы; смотрите также главу 8). У большинства нейронов много дендритов (мультиполярные нейроны), и если одни нейроны контактируют с другими через синапсы, то некоторые осуществляют эти контакты через дендритические отростки, или геммулы. Таким образом, основное назначение дендритов состоит в увеличении поверхности для синапсов, что позволяет интегрировать огромное количество сигналов, предназначенных телу нейрона. В противоположность этому аксон передает информацию от тела нейрона к нервной терминала и синапсам (см. главу 8). Несмотря на то что у каждого нейрона только один аксон, он разветвляется и имеет отростки. Эти ветви отходят вблизи тела нейронов, например чувствительных (псевдоуниполярные нейроны; см. главу 20), но чаше всего вблизи синапса, формируемого аксоном. Аксон отходит от аксонального бугорка, где начинается первый сегмент аксона. Это наиболее возбудимая часть нейрона, поскольку плотность натриевых каналов в этой области, где первоначально возникает потенциал действия, наиболее высока (см. главу 6). Все нейроны окружены лииидным бислоем (клеточной мембраной), где расположены белки, одни из которых образуют ионные каналы (см. главу 5), а другие рецепторы к специфическим веществам, высвобождаемым нейронами (см. главы 7 и 9), или же служат ионными насосами, которые перемещают ионы через мембраны в соответствии с электро-хнмическим градиентом, например Na -. К -ионный насос (см. главу 6). Поверхность мембраны аксона называется аксолеммой, а цитоплазма, которую она окружает, яксоплазмой. Ионные каналы аксо-леммы наделяют се способностью проводить потенциалы действия, а аксоплазма содержит микротрубочки, нейрофиламенты и митохондрии. Эти орга-пеллы не только отвечают за поддержание ионных фадиентов. но и обеспечивают транспорт и рециркуляцию белков из тела нейрона (и в меньшей степени к нему) к нервной терминали. Аксоплазмяти-ческийток, или аксональныйтранспорт, может быть медленным (примерно 1 мм/сут) или быстрым (100 400 мм/сут). Он необходим не только для поддержания нормальной активности нейрона и синапса, но и для жизнедеятельности и развития нейрона. Лк- сональный гок нарушается при некоторых пейроде-генеративпых заболеваниях (ем. главу 52). Многие аксоны покрыты слоем липидов, или миелиновой оболочкой, которая действует как электрический изолятор. Миелиновая оболочка влияет на способность аксона проводин, импульсы и способ ствует быстрому распространению потенциала действия без почери цс.кч i itcc ш импульса (см. глав) 8). Это достигается с помощью перехватов Ранвье — участков, в которых аксолемма содержит много ионных каналов (обычно № -каналов), непосредственно соприкасающихся с межклеточной жидкостью. Кроме того, из перехватов Ранвье отходят отростки аксона, которые называются коллатералями аксона. Миелиновая оболочка окружает аксон ниже аксонального холмика и заканчивается непосредственно вблизи нервной терминали, прежде чем та арбо-ризируется. Миелиновую оболочку формируют шванновские клетки в ПНС и олигодендроглиоциты в ЦНС (см. главу 4), при этом многие аксоны ЦНС окружает оболочка одного олигодендроглиоцита, тогда как в ПНС одна шванновская клетка приходится на один аксон. Синапс — это соединение между нейроном и другой клеткой. В ЦНС это другой нейрон. В ПНС это мышца, клетка железы или другого органа. В нервной системе обычно присутствуют химические синапсы, которые состоят из пресинаптической нервной терминали (концевой луковицы) и синаптичес-кой щели, которая отделяет нервную терминаль от постсинаптической мембраны и через которую выделяется химическое вещество, или нейротрансимит-тср (см. главу 7). Такой синапс обычно располагается между нейроном и дендритом другого нейрона (аксодендритический синапс), однако синапсы мо-iyr располагаться как в месте контакта между аксоном и постсинаптической клеткой, так и на теле нейрона (аксосоматические синапсы) и. реже, на пресинаптической нервной терминали (аксо-аксональные синапсы; см. главу Я). Некоторые синапсы ПНС не имеют таких свойств, но являются соединениями с малым сопротивлением (переходными соединениями) и называются апектрическими синапсами. Эти синапсы обеспечивают быстрое проведение потенциалов действия без какой-либо интеграции, что делает невозможным для популяции нейронов передавать импульсы вместе или синхронно (см. главы 7 и 53). 1 ибслы не....... ]..... 14 кн\ п<ч1\ляпнй нейронов при водит к развитию неврологических заболеваний. Такие заболевания, при которых гибель нейронов является первичным патологическим процессом, обсуждаются в главе 52. ГЛАВА 4. Клетки нервной системы II: неироглиальные клетки
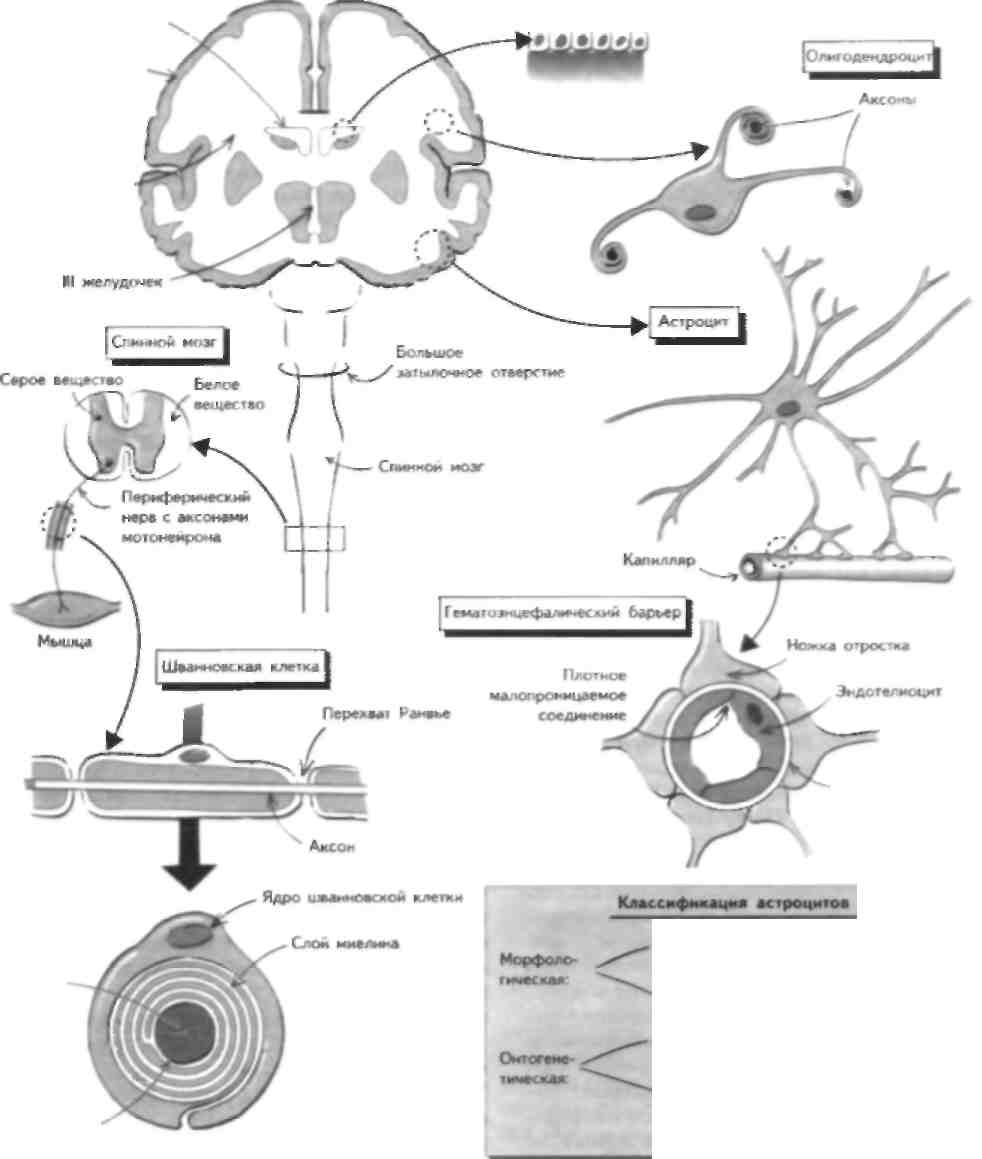
Боковой желудочек Кора мозга Полушарие головного мозга Эпендима Столбиковые щи mi alii jh. тм> _ клетки с ресничками
Выделяют 4 основных класса нейроглиальных клеток ЦНС: астроциты, олигодендроциты, эпенди-мальные и микроглиальные клетки, которые выполняют различные функции. В отличие отПНС, шван-новские клетки являются лишь частным примером iiciipoi.4iia.4i.них и очвечаюч за миелиничапию и ]Ч-генерацию аксонов. Астрощггы:»то маленькие звездчатые клетки. Их классифицируют по морфологическим и онтогенетическим признакам (см. рисунок). Они не являют- ся пассивными вспомогательными элементами ЦНС и выполняют многие важные ее функции: • Формируют вспомогательную сеть для нейронов • Поддерживают целостность гематоэнцефа.тическо-го барьера (ГЭБ), обеспечивая плотные соединения с малой проницаемостью между эндотелиоци-тами капилляров мозга (см. также главу 16). • Способны захватывать, хранить и высвобождать некоторые нейротрансмиттеры (например, глута-мат, у-аминомасляную кислоту — ГАМК) и поддерживают функцию химической нейротрансмиссии в ЦНС. • Могут захватывать избыточные ионы из экстра-целлюлярной жидкости, особенно калий. • Участвуют в процессе развития нейронов (см. главу 15), реагируют на повреждающее воздействие (см. главу 47), а также участвуют в судьбе клеток-предшественников нейронов в гиппокампе взрослого человека. Могут выполнять роль антигенпрезентирующих клеток в случаях, если ГЭБ и ЦНС повреждены. Наиболее распространенным заболеванием, связанным с поражением астроцитов, является астро-цитома. Опухоль сдавливает окружающую ткань ЦНС и приводит к развитию неврологического дефицита (с эпилептическими ириналками или без них), что зависит от се локализации. У взрослых ае-троцитома чаще всего локализуется в белом веществе полушарии мозга. Олигодендроциты отвечают за миелин и зацию нейронов ЦНС' и поэтому в большом количестве присутствуют в белом веществе. Каждый олнгодендро-инт образует миелиновую оболчкудля 3 50 волокон я также окружает многие другие волокна, формируя миелиновую оболочку. Кроме того, эти клетки выделяют ряд веществ, ингибирующих рост аксонов, едоватслыю, угнетают процессы регенерации нейронов в ЦНС взрослого человека (см. главу 47). Нарушение функции олигодендроцнтов приводит к демислипнзации ЦНС и отмечается при ряле заболеваний, например рассеянном склерозе (см. главу 54), тогда как при нарушении пролиферации олигодендроцнтов развиваются медленно растущие опу- холи (олиголендроглиомы), при которых характерны эпилептические припадки (см. главу 53). Эпендимальные клетки обеснечиваю'1 нормальную циркуляцию цереброспинальной жидкости (ЦСЖ), а также взаимодействую! с нстроцитами, формируя барьер, отграничивающий желудочки мола и ЦСЖ от нейроналыюго окружения. Они также выстилаю! центральный канал спинного мозга (см. главу 12). Эти клетки также называются эпен-димоцитами, поскольку другие эпендимальные клетки вырабатывают ЦСЖ (хороидные сплетения) и перемещают определенные вещества из ЦСЖ в кровь (танициты). Опухоли эпендимы (эпецаимомы и хороидные папилломы) возникают в желудочках, приводя к развитию гидроцефалии (см. главу 16), или в спинном мозге, где происходит локальная деструкция нервной ткани. Микрог. шальные клетки (не показаны на рисунке) являются тканевыми макрофагами мозга и находятся как в белом, так и в сером веществе ЦНС. По своей природе они являются фагоцитами и регулируют иммунный ответ в ЦНС (см. главу 54). Шванновские клетки находятся только в ПН С и отвечают за миелинизацию периферических нервов. окружая их аксоны миелиновой оболочкой. Завершенная миелиновая оболочка состоит из нескольких слоев мембраны шванновских клеток без цитоплазмы. В отличие от олигодендроцнтов, одна шван-новская клетка окружает аксон и образует миелин одного его участка между перехватами Ранвье. Кроме того, шванновские клетки участвуют в регенерации поврежденных аксонов, что отличает ИХ 01 пси тральных нейроглиальных клеток, которые подавляют процессы регенерации (см. главы 46 и 47). Гибель миелина (в отличие от гибели аксонов) происходит при ряде наследственных и воспалительных поли-нейропатнй, что при ноли i к л псф\ нмцш перифери ческих нервов (демиелинизнрующие невропатии; см. главы 6 и 55). Кроме того, известны доброкачественные опухоли шванновских клеток (шванно-мы), которые развиваются, в частности, при таком наследственном заболевании, как нейрофнброма-тоз I типа, связанном с нарушением функции гена-сунрессора опухолевого роста нейрофнбромина. Эти опухоли обычно протекают бессимптомно, од на ко если они образуются в ограниченном пространстве, то MOiyT вызывать славление окружающих нервных структур, например, в области мостомоз-жечкового угла или корешка спинномозгового нерва (см. главы 2 - 14 и 27). ГЛАВА 5. Ионные каналы i Вольтажмаисммый ионный канал Химический ионный канал
например, Ацх-катиоииый канал Место связывании Ацх (рецептор Ацх) Ионный канал, ассоциированный с рецептором
Инактиеированное состояние, канал закрыт
Нет ионного потока Ма' Селективный фильтр ?8 Вольтаж зависимый сенсор Субьединица — или ворота инактивации Приток натрия в клетку Na' Нет ионного потока Na*
Ионный канал это белковая макромолекула. которая располагается в биологической мембране и позволяет ионам переходить с одной стороны мембраны на другую. Ионы перемещаются через мембрану в направлении, определяемом электрохимическим градиентом. В общем, ионы стремятся из области более высокой их концентрации в облаем, более низкой, однако в присутствии вольтажзависимого градиента возможна ситуация, мри которой ионные потоки отсутствуют даже при наличии неравных концентраций ионов. Сам ионный канал может открываться и закрываться. Открытие его может быть связано с изменением заряда мембраны (например, при деполяризации или возникновении потенциала действия) или с взаимодействием химического вещества и рецептора в самом канале или поблизости от него. Выделяют два типа каналов вольтажзави-симые (вольтажчувствительные) и химические (ли-гандзависимые). Однако это разделение достаточно условно, поскольку многие вольтажзависимыс каналы могут быть активированы нейротрансмиперами и кальцием. Ьолсстого, некоторые каналы не открываются при изменении заряда мембраны или взаимодействии с химическими всществами-мсссснджс-рами, а реагируют только на давление и механическую силу (например, соматосснсорныс слуховые рецепторы; см. главы 2(1, 21 и 26). Наиболее важным свойством ионных каналов является возможность изменять электрическую возбудимость нейрона (см. главу 6). Ионные каналы находятся во всех частях нейрона, в меньшей степени в нейроглиальных клетках, а также в клетках неней-роналыюго происхождения. Все биологические мембраны, включая нейро-нальные, состоят из липидного бислоя, который характеризуется электрическим сопротивлением, т. е. ионы не могут просто проникнуть через него. Поэтому для облегчения проникновения ионов через мембраны существую! так начинаемые поры (ионные каналы) на липидной мембране или переносчики, которые собирают ионы с одной стороны мембраны и переносят их через нее, а потом высвобождают ионы. В нейронах скорость переноса ионов, необходимая для передачи импульса, слишком высока для какой-либо системы переносчиков, поэтому в нейронах и существуют ионные каналы (или поры) для переноса ионов через мембраны. Основные свойства ионных каналов таковы: • Они состоят из нескольких белковых субъединиц, которые локализуются трансмембран но и позволяют ионам переходить с одной стороны мембраны на другую, — трансмембранные поры. • Канал сформирован таким образом, что он может открываться и закрываться, при этом данный процесс имеет несколько промежуточных ступеней. • Канал может открываться в ответ на специфические стимулы. Большинство каналов реагирует на изменение заряда мембраны и поэтому открывается в ответ на деполяризационный заряд, т. е. на заряд, меняющийся на мембране в покое от —70 до 80 мВ к более отрицательной величине. В противоположность этому некоторые ионные каналы, особенно синаптические, не открываются в ответ на изменение заряда мембраны, а открываются в ответ на взаимодействие с химическими веществами (например, ацетилхолином Ацх). Эти каналы имеют в своей сфукзурс рецептор для этого вещества, и связывание его с этим рецептором приводит к открытию канала. Однако многие ионные каналы обладают как вольтажзависимыми, так и химическими сенсорами и наличие в интранеллюляр-ном пространстве иона или вторичного мсссепджс-ра — циклическою аденозинмонофосфата (цЛМФ) приводит к модуляции трансмембранного ионного потока, вызванною вольтажзависнмым процессом. Активация вольтажчувствитсльного или химического рецептора приводит к открытию ворот каша, через которые осуществляются ионные потоки. Зачем капал закрывается под воздействием дезактивации (процесс, обрачпий открытию канала) или инактивации, что предполагает участие вторичных ворот, через которые но каналу ионы поступают медленнее, чем выходят по активированным воротам, и таким образом существует временной промежуток, когда в канале нет ворот и ионы не могут через него пройти. Ионные потоки через канал могут быть сатектив-ными и неселективнымн. Если канал селективен, он обеспечивает прохождение только определенных ионов, что достигается посредством фильтра. В основе селективного фильтра лежат термодинамические свойства. Такому каналу присваивается назва- ние, например «натриевый канал». Однако многие каналы не являются селективными и пропускают различные виды ионов с одноименным зарядом, например Лцх-катионный канал. Ионный канал характеризуется определенными физическими свойствами. Перемещение ионов но каналу называется потоком, тогда как термином «проводимость» определяют рсципрокное сопротивление (поток/вольтаж). Проводимость это степень доступности перемещения иона по каналу. Прони-цяемость, с другой стороны, определяется как скорость перемещения вещества или иона через мембрану при определенной разности концентраций. Существует множество разновидностей ионных каналов даже в пределах одного семейства специфически ионных каналов, например калиевых. Количество и тип ионных каналов определяют характеристики клеточного ответа. Например, нейроны характеризуются скоростью генерации потенциала действия и ответа на синаптические импульсы (см. главы 6—8, 45 и 53). Дата добавления: 2015-02-06 | Просмотры: 2002 | Нарушение авторских прав |

 например, натриевый канал
например, натриевый канал