|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Нервы (II и IV пары ЧМН) студенистый материал, называемый
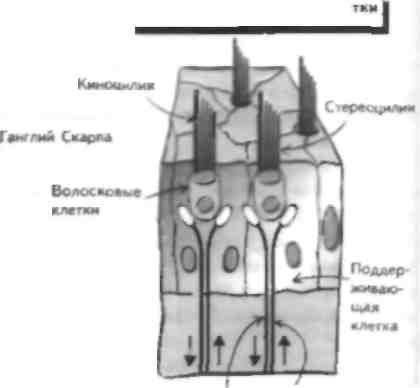
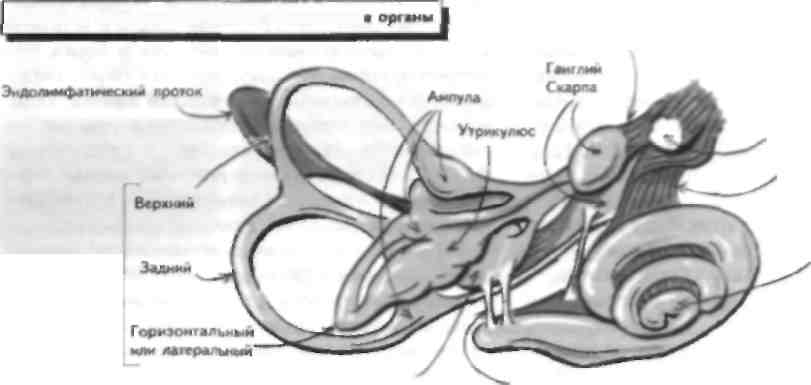
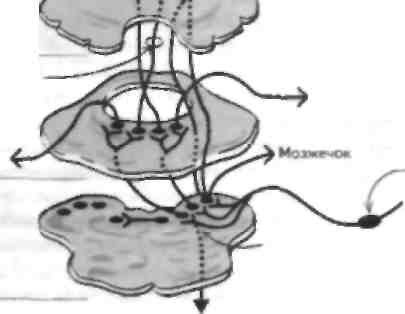
Вестибулярная система отвечает за поддержание равновесия, реализацию постуральных рефлексов и регуляцию движений глаз. Она состоит из периферического воспринимающего компонента, который проецируемся веч вол мозга (включая глазодвигательные ядра), а сviтуда в |аламус и на сенсорную КОру, а также на мозжечок и в спинной мозг. Нарушение функции этой системы приводит к развитию голо- вокружснии, тошноты и рвоты, иногда в сочетании с глазодвигательными нарушениями (обычно нистагмом; см. главу 40) и неустойчивостью при ходьбе. У больного в коме исследование вестибулярного анализатора лает важную информацию о степени целостности ствола мозга. Вестибулярное восприятие Периферический воспринимающий компонент вестибулярного анализатора состоит из лабиринта, который в свою очередь включает два отолитовых органа (утрикулюс и енккулюс), а также ампулы полукружных каналов. Отолитовые органы воспринимают информацию о положении головы и линейном ускорении, тогда как полукружные каналы о вращательном (угловом) ускорении. Волосковые клетки находятся в обоих отолитовых органах и сходны по структуре с волосковыми клетками улитки (см. главы 19 и 26). Как и в улитке, смещение стереоцилий в сторону киноцилии деполяризует клетку и позволяет нейротрансмиттеру высвободиться из волосковой клетки, что приводит к активации ассоциированного афферентного волокна. Когда стереопилий смешаются в противоположном направлении, этого не происходит. Движения ресничек связаны с вращением головы (ампулярные рецепторы в полукружных каналах) и ускорением при ее наклоне (отолитовые органы утрикулюса), так как движение головы вызывает перемещение эндо-лимфы, которая в свою очередь омывает волосковые клетки и вызывает их движение, изменяя конфигурацию стереопилий. Спонтанная активность афферентных волокон высокая и отражает спонтанное вытекание нейро-трапемиттера из волосковой клетки в синапс. Гн-периоляризация волосковой клетки приводит к уменьшению афферентного разряда, тогда как деполяризация связана с увеличением импульсацни. Эфферентные волокна ствола мозга, закапчивают неся па волосковых клетках, могут изменить чувствительность рецепторов воспринимающего органа. Периферические расстройства вестибулярного анализатора Поражение периферической вестибулярной системы встречается часто. Примером такого поражения является доброкачественное позиционное головокружение, которое часто возникает после травмы пли инфекции вестибулярного аппарата, когда отолитовые кристаллы скапливаются в заднем полукружном канале. Это заболевание характеризуется приступами головокружения, тошнотой, рвотой и атаксией при поворотах головы в стороны, например при укладывании головы па подушку и поворотах в постели. Эти симптомы связаны с перемещением эндолим-фы в заднем полукружном канале вслед за перемещением отолитовых рецепторов. Симптомы можно устранить, если совершить несколько поворотов головы, при которых отолитовые рецепторы упадут в полукружный канал и ампулу. Вирусные инфекции вестибулярного аппарата встречаются часто (лабн-ринтит) и MOiyr значительно ухудшать качество жизни больных, привили к развитию тяжелых приступов тошноты п рвоты. Инфекции вестибулярного аппарата обычно самопроизвольно прекращаются. Двустороннее поражение вестибулярного аппарата может привести к оецнллопсин, которая заключается в неспособности фиксировать взгляд на объектах при движениях головы (см. главу 40). В противоположность этому раздражение вестибулярной системы, например, при укачивании приводит к головокружению, тошноте, рвоте и тахикардии, вызванными рассогласованностью вестибулярной и зрительной информации. Вестибулярную функцию можно проверить, вводя воду в наружный слуховой проход (калорическая проба). Когда в положении сидя в ухо вводят теплую воду и пациент наклоняет голову назад на 60°. возникает нистагм в сторону раздражаемого лабиринта. Введение холодной воды вызывает нистагм в противоположную сторону. Эти изменения отражают перепады температуры эндолимфы, которые дают эффект, сходный с вращением головы в сторону, противоположную от раздражаемого лабиринта. Центральная часть вестибулярного анализатора и вестибулярные рефлексы Тела нейронов, дающих афферентные вестибулярные волокна в составе VIII пары ЧМП, лежат в вестибулярном ганглии Скарна и заканчиваются в одном из 4 вестибулярных ядер в продолговатом мопс, которые также получают импульсацию от рецепторов мышц шеи и зрительного анализатора. Вестибулярные ядра проецируются в спинной мозг (см. главы 12, 32 и 34), противоположные вестибулярные ядра, ядра глазодвигательных нервов, а также в таламусы. Некоторые из этих структур имеют важное значение дня реализации рефлекторных движений глаз, таких, как способность фиксировать взгляд на предмете при движении головы окулове-стибулярнып рефлекс (ОВР; см. главы 40 п 47). Дру-гас проекции вестибулярных ядер играют важную роль в поддержании позы и ходьбе. Вестибулярные волокна заканчиваются в первичной соматосенсорной коре (ПС'СК) и задней теменной коре (см. главу 30). Калорические пробы позволяют уточнить степень целостности вестибулярного аппарата и его стволовых связен, поэтому они Moiyi ими. исполь зованы у больных в коме для уточнения степени поражения ствола. Менее тяжелые поражения вестибулярного аппарата Moiy i возникать при других забо левапиях, например рассеянном склерозе (см. главу 54) и инсультах (см. главу 17). Иногда в патологические принесем вовлекаются п другие структуры. ГЛАВА 29. Обоняние и вкус
Синапс > Афферентные МЦВ '!
Обонятельный, или I черепно-мозговой нерв (ЧМН) содержит наибольшее количество волокон по сравнению с другими чувствительными нер- вами, проецирующимися в ЦНС. Вкусовые ощущения проводятся в ЦНС волокнами VII, IX и X пар ЧМН. Обоняние Вкус
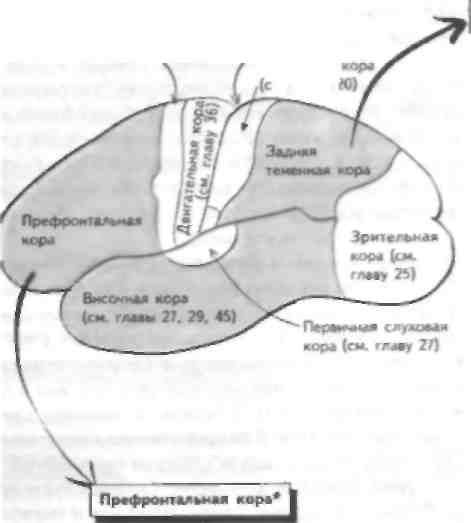
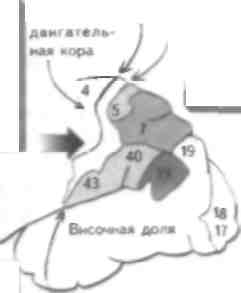
Обонятельный анализатор предназначен для различения большого количества запахов, что возможно благодаря наличию тысяч обонятельных рецепторов. Эти репепторы представляют собой апикальные дендриты обонятельных рецепторных клеток, которые проецируются в ЦНС через продырявленную пластинку решетчатой кости в верхних отделах носовой полости на обонятельную луковицу. Обонятельный стимул или запах связывается с обонятельными рецепторами и деполяризует их (см. главу 19), что при достаточном уровне стимуляции приводит к генерации потенциала действия в теле нейрона и проведению этого импульса по аксонам обонятельного нерва в обонятельную луковицу. Обонятельный нерв проходит через крышу носовой полости через продырявленную пластинку решетчатой кости. Поражение этой структуры (например, при черепно-мозговой травме) может привести к повреждению аксонов обонятельного нерва и утрате обоняния — аносмии. Однако чаще всего нарушения обоняния возникают при инфекции или воспалении слизистой оболочки полости носа. Лксоны обонятельных рецепторов переключаются на нейроны обонятельной луковицы, которая залегает в базальных отделах лобной доли. Поражение этой структуры возникает при базальных ме-нингиомах и приводит к развитию односторонней апосмии. Обонятельная луковица содержит комплекс клеток. Лксоны обонятельного нерва образуют синапсы с апикальными депдритамн митральных и в меньшей степени пучковых клеток, аксоны которых проецируются в виде обонятельного тракта. Обонятельная луковица содержит ряд тормозных ингерпейро-нов (гранулярных и перигломеруляриых клеток), которые модифицируют обонятельные стимулы, проходящие через луковицу. Обонятельный тракт проецируется па височную долю и образует синапсы с корой грушевидной извилины п лимбичеекой системой, которая проецируется на гипоталамус. Эта проекция важна для реализации поведенческого компонента обоняния, который наиболее развит у животных. У человека поражение этих структур редко приводит к развитию чистой апосмии, а активация этих отделов ЦНС вызывает височную эпилепсию (см. главу 53), при которой возникают нарушения восприятия запахов (например, обонятельные галлюцинации). Обонятельная система дает проекции на таламус и через обонятельный бугорок на медиодорсальное ядро, которое проецируется на префронтальную кору. Роль этого пути неясна. Вкусовые рецепторы расположены на языке. Они собраны зам в пиле грибовидных сосочков и cibo НОВЫХ клеток; последние делятся и замещают вкусовые рецензоры при их повреждении. Апикальная поверхность вкусового рецептора содержи! микроворсинки, покрытые слизью, которая вырабатывается соседними бокаловидными клетками. Каждый продукт достигает вкусового рецептора; гидрофильные вещества растворяются в слюне, алипофильные - в слизи. Вкус имеет несколько модальностей: соленый, кислый, сладкий и горький. Эти виды вкусовой чувствительности соответствуют определенным типам восприятия этих ощущений. Недавно был открыт новый вкус (умами). Соленые импульсы вызывают непосредственную деполяризацию вкусовых рецепторов при прохождении натрия по амилоридчувствительному каналу апикальной мембраны. Деполяризация приводит к выс-вобожаению нейротрансмиттера из базальной части клетки, который активирует афферентные волокна соответствующего ЧМН. В отличие от соленых, кислые вкусовые стимулы блокируют вольтажзависимые протонные каналы. Сладкие стимулы связываются с рецептором, который активирует G-белок густиду-цин, который активирует аденилатциклазу и вырабатывается цЛМФ. Повышение концентрации цЛМФ приводи! какишацнп прозеинкипазы, которая (рос форшшруется и закрывает базолатеральные калиевые каналы и таким образом деполяризует рецептор. Горькие стимулы переключаются на рецептор посред-езном ак'шванпн О-белка. При акзпвапии |уетидупн-на активируется цЛМФ-фосфодиэстераза, которая снижает уровень цЛМФ (и фосфорилироиания прозеиикииазы), чю нриводиз к открытию Оазолатсраль-ных кальциевых каналов и высвобождению нейротрансмиттера. Другой способ передачи сладких и горьких стимулов заключается в активации фосфо-,iini:rn,I С и iip<vi\kniiii ииозизолтрифосфаза (И I Ф). а также диапнлглиперола (ДАТ), которые способствуют высвобождению кальция из внутренних запасов рецептора Увеличение концентрации кальция при водит к высвобождению нейрозрансмизтера. Рецепторы передают информацию по барабанной струне (передние 2/3 языка) и языкоглоточному нерву (задняя треть языка) к ядру одиночного пути в продолговатом мозге (см. главы 13 и 14). Эта структура проецируется рострально через таламус в ПССК и кору островковой извилины с последующей проекцией на гипоталамус и миндалевидное тело. У больных височной эпилепсией наблюдается аура в виде обонятельных галлюцинации, в возникновении которых принимает участие проекция на гипоталамус и миндалевидное тело (см. главу 53). ГЛАВА 30. Ассоциативная кора: задняя теменная и префронтальная кора
Корковые отделы: от все» отделов коры, ио особенно от теменно-затылочных Дата добавления: 2015-02-06 | Просмотры: 1255 | Нарушение авторских прав |
 купулой
купулой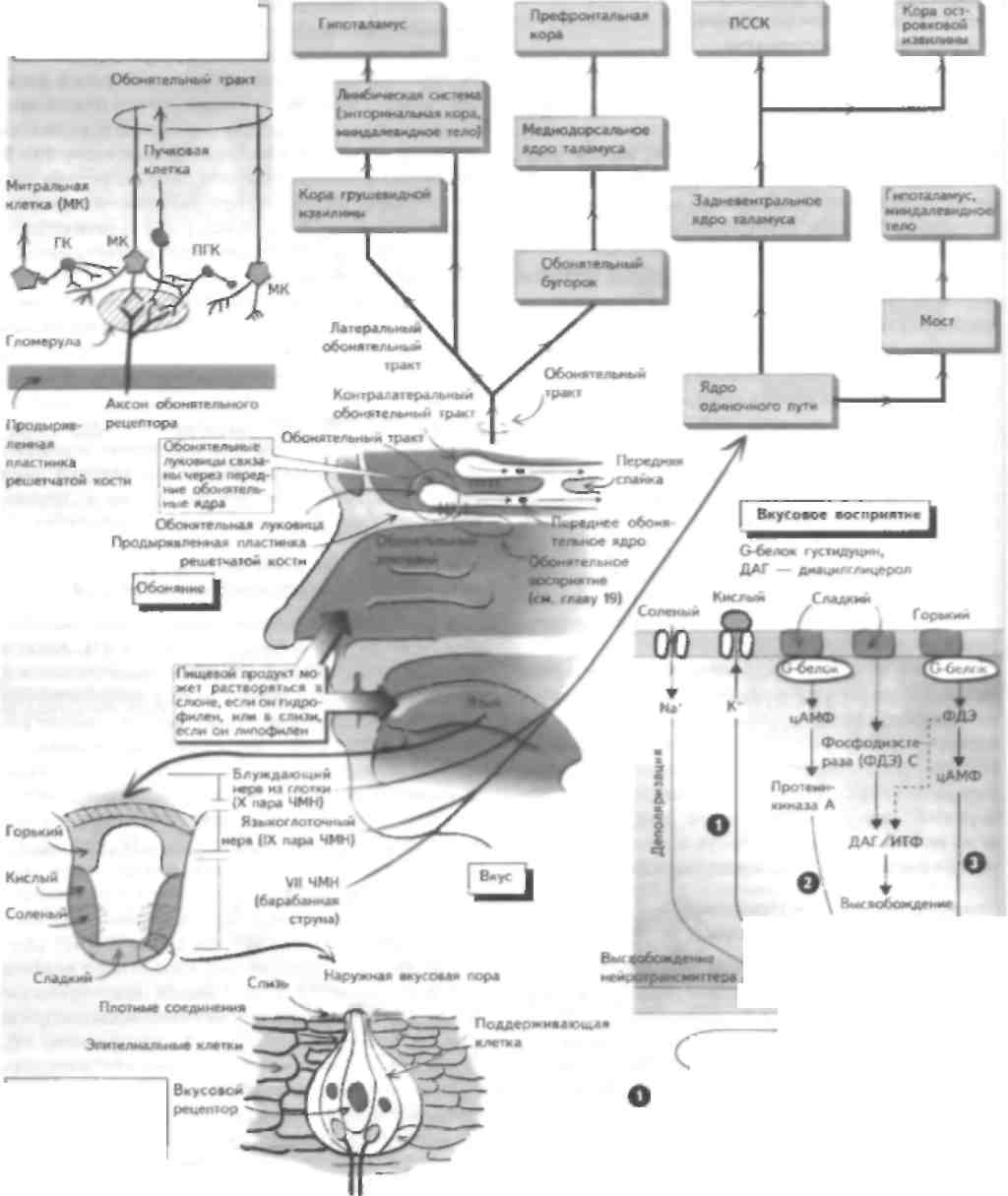
 Афферентные волокна
Афферентные волокна