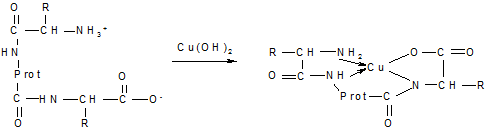|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Строение и свойства аминокислот и пептидовАминокислоты — соединения, в молекулах которых одновременно присутствуют амино- и карбоксильные группы. Природные α-аминокислоты
являются биологически активными соединениями; их классификация приведена в табл. 16. Таблица 16 Классификация природных α-аминокислот
Строение аминокислот. Все α-аминокислоты* можно рассматривать как результат замены атома водорода в простейшей а-аминокислоте — глицине — на тот или иной радикал R. Таким образом, в соответствии с природой радикала R, называемого боковой цепью, -аминокислоты подразделяют на 4 группы, отличающиеся гидрофильностью или гидрофобностью боковых цепей, а также способностью боковой цепи проявлять кислотные или основные свойства (см. табл. 16). Стереохимия природных α-аминокислот характеризуется тем, что все они кроме глицина имеют асимметрический атом углерода (атом, связанный и с амино-, и с карбоксильной группой), конфигурация которого может быть отождествлена с конфигурацией L-глицеринового альдегида путем цепи химических превращений:
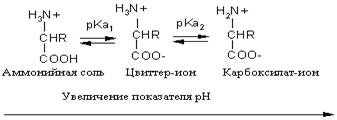
* За исключением нейтральной гидрофобной аминокислоты пролина (L-пирролидин-α-карбоновой кислоты). При этом превращения либо не должны затрагивать хиральный центр, либо реакции должны протекать строго стереоспецифично. Следовательно, все природные α-аминокислоты являются L-энантиомерами. Конфигурация асимметрического центра аминокислот определяет биологические свойства как самих аминокислот, так и олиго- и полимерных соединений, мономерами которых служат остатки аминокислот (эти соединения называют пептидами). Свойства аминокислот. Аминокислоты представляют собой бесцветные кристаллические вещества с довольно высокими температурами плавления (более 230 °С). Большинство кислот хорошо растворимы в воде и практически не растворимы в спирте и диэтиловом эфире, что указывает на солеобразный характер этих веществ. Специфическая растворимость аминокислот обусловлена наличием в молекуле одновременно аминогруппы (имеющей основный характер) и карбоксильной группы (характеризующейся кислотными свойствами), благодаря чему аминокислоты принадлежат к амфотерным электролитам (амфолитам). В водных растворах и твердом состоянии аминокислоты существуют только в виде внутренних солей — цвиттер-ионов. Кислотно-основное равновесие для аминокислоты может быть описано следующим образом:
Если к раствору аминокислоты приложено электрическое поле, то в зависимости от показателя рН раствора ионы аминокислоты будут перемещаться по-разному: в кислой среде при рН < 7 аммонийные ионы аминокислот перемещаются к отрицательному полюсу (катоду), а в щелочной среде при рН > 7 карбоксилат-ионы — к положительному полюсу (аноду). Значение рН, при котором молекула аминокислоты электронейтральна, называют изоэлектрической точкой и обозначают р I. При значении рН, равном показателю р I, молекула аминокислоты в электрическом поле не перемещается. Изоэлектрическую точку определяют по соотношению p I =0.5(p K a1+p K a2)
Реакции с участием только аминогруппы. Наличие в молекуле одновременно амино- и карбоксильной группы отражается и на поведении аминокислот в тех реакциях, в которых участвует только одна из двух функциональных групп. Аминогруппа, которая в аминах проявляет себя как нуклеофил, в биполярном ионе полностью лишена нуклеофильности из-за протонирования, поэтому ни реакция алкилирования по Гофману, ни ацилирование, свойственные аминам, не имеют места в случае аминокислот. Эти реакции могут происходить только при условии предварительного депротонирования аминогруппы, что достигается использованием реакционной среды с высоким значением рН, при которых цвиттер-ион полностью превращен в карбоксилат-анион. Рассмотрим основные реакции с участием только аминогруппы аминокислот. 1. Алкилирование осуществляют, действуя на полученные соли аминокислот алкилгалогенидами в присутствии оснований (как органических, так и неорганических). 2. Ацилирование также требует предварительного превращения цвиттер-иона в карбоксилат-анион и успешно протекает при наличии в реакционной среде эквивалента основания (основание необходимо для связывания выделяющегося при ацилировании кислого продукта — галогеноводорода или карбоновой кислоты):
или
и далее
3. Образование оснований Шиффа (как типичная реакция аминов) свойственно и аминокислотам; наиболее часто используют реакции аминокислот с бензальдегидом:
На образовании оснований Шиффа основана реакция идентификации аминокислот, известная как «нингидриновая проба», широко применяемая для визуализации зон аминокислот (возникает интенсивное сине-фиолетовое окрашивание) при их хроматографическом и электрофоретическом разделении, а также для количественного определения содержания аминокислот в растворах:
Эта реакция лежит в основе метода определения содержания азота и количества аминогрупп в аминокислотах (метод Ван-Слайка). Биосинтез аминокислот. Все природные α-аминокислоты делят на незаменимые (валин, гистидин, изолейцин, лейцин, лизин, метионин, треонин, триптофан, фенилаланин), которые поступают в организм только из внешней среды, и заменимые, синтез которых происходит в организме. Биосинтез α-аминокислот может происходить на основе не аминокислот, например по реакции восстановления
Реакция стереоспецифична вследствие стереоспецифичности НАДН. В качестве исходных веществ при биосинтезе аминокислот могут выступать другие аминокислоты. Например, реакция трансаминирования (переаминирования) является основной при синтезе α-аминокислот в организме:
Катализаторами и участниками этого процесса являются ферменты (аминотрансферазы) и кофермент пиридоксальфосфат, который служит переносчиком аминогруппы. Пептиды. Амино- и карбоксильные группы аминокислот могут реагировать друг с другом, даже если они находятся в одной молекуле. Еще более реальным является образование межмолекулярной амидной связи. Амиды, образовавшиеся в результате взаимодействия некоторого числа аминокислот, называют пептидами. В зависимости от числа аминокислотных остатков различают ди-, три-, тетра-, пентапептиды и т.д. При этом пептиды молекулярной массой не более 10 000 называют олигопептидами; молекулярной массой более 10 000 — полипептидами, или белками. Амидные связи в составе пептидов называют пептидными. Пептидная группировка характеризуется рядом свойств. 1. Пептидная группировка имеет жесткую планарную структуру, т. е. все атомы, входящие в нее, располагаются в одной плоскости. 2. Атомы кислорода и водорода пептидной группировки природных пептидов и белков находятся в транс -положении по отношению к связи С—N, так как при транс -конфигурации заместителей боковые цепи оказываются наиболее удалены друг от друга, что важно для стабилизации структуры белковой молекулы. 3. Пептидная группа представляет собой трехцентровую ρ, π-сопряженную систему, которая образуется вследствие делокализации электронной плотности между атомами кислорода, углерода и азота. Длины связей С—О и С—N оказываются практически одинаковыми. 4. Пептидная связь устойчива при температуре 310 К в средах, близких к нейтральной (физиологические условия). В кислой и щелочной средах связь подвергается гидролизу. В условиях организма гидролиз происходит ферментативно. 5. Дополнительные, как правило, нековалентные связи между пептидной группой и боковыми цепями обусловливают существование различных конформаций белковой молекулы. Например, внутримолекулярные водородные связи 6. Пептидная группировка может существовать в двух резонансных формах (кетонной и енольной):
Эти свойства пептидной группировки определяют строение полипептидной цепи:
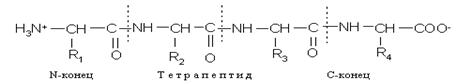
Полипептидная цепь состоит из регулярно повторяющихся участков, образующих остов молекулы, и вариабельных участков — боковых радикалов аминокислотных остатков. Началом полипептидной цепи считают конец, несущий свободную аминогруппу (N-конец), а заканчивается полипептидная цепь свободной карбоксильной группой (С-конец).
Как правило, при изображении формулы пептида N-конец располагают слева, а С-конец — справа:
Называют пептид, последовательно перечисляя, начиная с N-конца, названия аминокислот, входящих в пептид; при этом суффикс «ин» заменяют на суффикс «ил» для всех аминокислот, кроме С-концевой. Для описания строения пептидов применяют не традиционные структурные формулы, а сокращенные обозначения, позволяющие сделать запись более компактной. Понятие «строение пептида» (равно как и «структура белка») включает в себя следующие характеристики: 1) общее число аминокислотных остатков; 2) перечень аминокислот, входящих в состав пептида, и указание количества аминокислотных остатков каждого вида (этот параметр называют аминокислотным составом пептида или белка ); 3) последовательность связывания аминокислот друг с другом (этот параметр называют аминокислотной последовательностью; он отражает так называемую первичную структуру пептида или белка); последовательность записывают слева направо от N-конца к С-концу. Особенности растворения полимеров. Размеры макромолекул высокомолекулярных соединений (ВМС) соизмеримы с размерами коллоидных частиц, что обусловливает общность ряда свойств, характерных для коллоидных растворов и растворов ВМС. К таким свойствам относят малую скорость диффузии растворенных частиц, неспособность их проникать через мембраны, эффект Фарадея—Тиндаля и др. Однако растворы ВМС являются истинными, поскольку удовлетворяют основным критериям истинных растворов: самопроизвольность образования, гомогенность, термодинамическая устойчивость, равновесность. Равновесие в растворах ВМС устанавливается медленнее, чем в истинных растворах, и растворению, как правило, предшествует набухание. Набуханием называют самопроизвольный процесс односторонней диффузии низкомолекулярного растворителя в полимер, сопровождающийся увеличением объема и массы ВМС. Различают неограниченное и ограниченное набухание. В первом случае полимер поглощает жидкость, а потом при той же температуре постепенно переходит в раствор. Пример неограниченного набухания — растворение желатина или крахмала в горячей воде. При ограниченном набухании процесс практически останавливается на стадии образования гетерогенной системы, состоящей из двух фаз: набухший полимер и низкомолекулярный растворитель. В этом случае равновесной системой является гель. Пример ограниченного набухания — набухание желатина или крахмала в холодной воде, набухание резины в бензоле. Тип набухания зависит от гибкости полимерной цепи: чем более гибкой является полимерная цепь, тем больше степень набухания и тем выше вероятность образования раствора. Количественно способность полимера набухать в тех или иных растворителях характеризуют степенью набухания:
где V, V0 — объем образца полимера до и после набухания соответственно. На практике для оценки способности растворителя растворять или вызывать набухание того или иного полимера обычно руководствуются эмпирическим правилом: подобное растворяется в подобном (неполярные полимеры растворяются в неполярных растворителях, а полярные — в полярных). Для амфотерных полиэлектролитов степень набухания зависит от показателя рН среды. Так, белки в изоэлектрическом состоянии имеют минимальные значения степени гидратации, набухания, растворимости. Зависимость степени набухания белка от значения рН среды выражается кривой с двумя максимумами и одним минимумом, который соответствует изоэлектрической точке. Качественные реакции. Для идентификации некоторых пептидов и белков используют так называемые «цветные реакции». Наиболее универсальная реакция на пептидную группу — появление красно-фиолетовой окраски при добавлении к раствору белка ионов меди(П) в щелочной среде (биуретовая реакция):
Реакция на остатки ароматических аминокислот — тирозина и фенилаланина — появление желтой окраски при обработке раствора белка концентрированной азотной кислотой (ксантопротеиновая реакция):
Серасодержащие белки дают черное окрашивание при нагревании с раствором ацетата свинца(II) в щелочной среде (реакция Фоля):
Цветные реакции на белки. Для белковых веществ характерен ряд цветных реакций, которыми обычно пользуются для обнаружения белков в биологических объектах. Важнейшие них следующие. Биуретовая реакция — сине-фиолетовое или красно-фиолетовое окрашивание при прибавлении к водному раствору белка щелочи и медного купороса. Эта реакция зависит от наличия в белках пептидных связей СО—NH. Ксантопротеиновая реакция — появление желтого окрашивания под действием концентрированной азотной кислоты. Эта реакция характеризует наличие в белках цикличес- ких аминокислот (тирозина, фенилаланина, триптофана). Реакция Миллона — розовое или красное окрашивание при нагревании белков с реактивом, состоящим из смеси азотнокислых и азотистокислых солей ртути в азотной кислоте. Реакция связана с наличием в белке фенильной группы тирозина. Реакция Адамкевича — сине-фиолетовое окрашивание при прибавлении к белку раствора глиоксиловой кислоты в серной кислоте. Этот цвет обусловлен конденсацией альдегидной группы глиоксиловой кислоты с индольным кольцом триптофана. Реакция Сакагуши — малиново-красное окрашивание при обработке белка сначала гипохлоритом натрия, а затем раствором β-нафтола. Эта реакция указывает на присутствие белке гуанидиновой группы аргинина.
Дата добавления: 2015-02-06 | Просмотры: 2140 | Нарушение авторских прав |









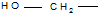

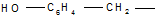






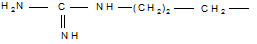



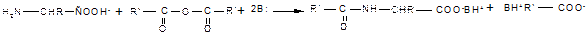


 4. Дезаминирование аминокислот, как и всякого первичного амина, протекает при действии на аминокислоты азотистой кислоты:
4. Дезаминирование аминокислот, как и всякого первичного амина, протекает при действии на аминокислоты азотистой кислоты:
 -кетокислот под действием НАДН:
-кетокислот под действием НАДН:

 стабилизируют вторичную структуру белка.
стабилизируют вторичную структуру белка.