|
АкушерствоАнатомияАнестезиологияВакцинопрофилактикаВалеологияВетеринарияГигиенаЗаболеванияИммунологияКардиологияНеврологияНефрологияОнкологияОториноларингологияОфтальмологияПаразитологияПедиатрияПервая помощьПсихиатрияПульмонологияРеанимацияРевматологияСтоматологияТерапияТоксикологияТравматологияУрологияФармакологияФармацевтикаФизиотерапияФтизиатрияХирургияЭндокринологияЭпидемиология |
Развитие, структура и функция яичников
Эмбриология
В течение 3-й недели беременности из эндодермальной выстилки желточного мешка на каудальном конце эмбриона образуются примордиальные зародышевые клетки. На 5-й неделе беременности они мигрируют к половому гребешку, расположенному около мезонефрической почки, и претерпевают митотическое деление. До 7-й недели внутриутробной жизни гонады находятся в недифференцированном состоянии, а несколько позже удается отличить примитивный яичник от яичка (см. гл. 333). Между 8-й и 10-й неделями беременности в яичнике начинают образовываться эстрогены, а на 10—11-й неделе отдельные оогонии в развивающемся корковом слое яичника превращаются в первичные ооциты. В яичнике содержится конечное число зародышевых клеток; максимум оогоний (около 7 млн) формируется к 5—6-му месяцу беременности. Затем число зародышевых клеток начинает уменьшаться в результате атрезии; к моменту родов их остается только 1 млн, к менархе — 400 тыс., а после менопаузы — всего несколько штук. Для нормального созревания яичников необходимы две Х-хромосомы; у индивидуумов с кариотипом 45, Х яичники, хотя и развиваются, но скорость атрезии возрастает настолько, что к моменту родов на их месте остаются лишь фиброзные тяжи (см. гл. 333). По окончании пролиферации оогоний начинается мейоз, продолжающийся до завершения стадии диплотены первого мейотического деления; затем ситуация остается стабильной до начала овуляции в период полового созревания. В течение 5-го месяца внутриутробной жизни формируется примордиальный фолликул, содержащий первичный ооцит, остановившийся в мейозе, тонкий слой гранулезных клеток и базальную мембрану, которая отделяет фолликул от окружающих тканей стромы (интерстициальной ткани).
Созревание в пубертатном периоде
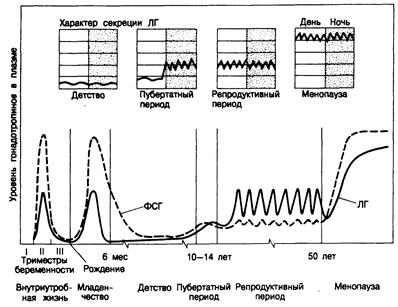
Окончательно фолликулы яичников созревают в пубертатном периоде. Развитие фолликула регулируют два основных гормона — гипофизарные гонадотропины фолликулостимулирующий гормон (ФСГ) и лютеинизирующий гормон (ЛГ) (рис. 331 -1). В течение II триместра внутриутробной жизни содержание гонадотропинов в плазме соответствует таковому в менопаузе. Этот пик гонадотропинов может служить причиной одновременного пика репликации ооцитов. Гипоталамо-гипофизарная ось (так называемый гонадостат) созревает и после II триместра становится чувствительной к действию стероидных гормо-
Рис. 331-1. Характер секреции гонадотропинов в разные периоды жизни женщины. ФСГ—фолликулостимулирующий гормон, ЛГ—лютеинизирующий гормон. Характер секреции ЛГ в часы бодрствования (светлая зона) и ночью (пунктирная зона) в разные периоды показан на верхней части рисунка. (По С. Faiman et al.)
нов крови, особенно плацентарным эстрогенам и прогестерону, по механизму отрицательной обратной связи. Поэтому содержание гонадотропинов в крови снижается и к моменту рождения становится практически неопределимым. В результате отделения плода от плаценты у новорожденных происходят снижение уровня эстрогенов и прогестерона и одновременное ответное повышение секреции гонадотропинов, сохраняющееся в течение первых месяцев жизни. Гипоталамо-гипофизарная система продолжает созревать, и гонадостат становится чувствительным к регуляции по механизму отрицательной обратной связи со стороны низкого уровня стероидных гормонов в крови. В результате содержание гонадотропинов в плазме вновь падает. С приближением пубертатного возраста чувствительность гонадостата снижается, а секреция ФСГ и ЛГ увеличивается, возможно, в результате повышения продукции гипоталамусом рилизинг-гормоналютеинизирующего гормона (ЛГРГ) (см. гл. 321). Затем секреция ЛГ во сне приобретает пульсирующий характер — первый этап формирования импульсного характера секреи(ии гонадотропинов (см. рис. 331-1). Последующее повышение секреции эстрогенов уже по механизму положительной обратной связи усиливает импульсную секрецию ЛГ и приводит в конце концов к овуляции и менархе, затем средняя концентрация гонадотропинов в плазме достигает уровня, характерного для взрослых. Содержание их в дневные и ночные часы выравнивается. После наступления менопаузы уровень гонадотропинов в плазме возрастает, через 5—10 лет достигает плато и остается довольно постоянным до 80—90-летнего возраста, а затем уменьшается. Хотя функция яичников регулируется в основном ЛГ и ФСГ, эти железы содержат и рецепторы пролактина и ЛГРГ. В яичниках человека in vitro оба эти гормона ингибируют стероидогенез, что позволяет предполагать их роль в патофизиологии этих желез. В период полового созревания чувствительность гипоталамо-гипофизарных центров к стероидным гормонам крови снижается, возрастает секреция ЛГРГ гипоталамусом, гонадотропинов гипофизом, эстрогенов яичниками и появляются анатомические изменения, свойственные этому периоду. Первые вторичные половые признаки у девочек становятся заметными в 10—11 лет. Начинают увеличиваться грудные железы (телархе), появляются волосы на лобке (пубархе), а затем в подмышечных впадинах (адренархе). Считают, что рост волос на лобке и под мышками обусловлен повышением секреции надпочечниковых андрогенов, начинающимся в 6—8-летнем возрасте. Происходит бурное ускорение роста; максимум его скорости приходится в среднем на 12-летний возраст. О половом созревании свидетельствует начало регулярных циклических менструаций. От начала развития грудных желез до наступления менструаций (менархе) проходит в среднем 2 года. Возраст, когда у девочек наступает первая менструация, варьирует и отчасти определяется не только генетическими факторами и общим состоянием здоровья, но и социально-экономическими условиями. Полагают, что в США за последние 100 лег средний возраст менархе снижался со скоростью 3—4 мес за 10 лет и в настоящее время он составляет примерно 13 лет. Такое снижение объясняют улучшением питания населения. Причем масса тела в этот срок должна быть около 48 кг или имелось нужное соотношение массы, содержания воды и жира в организме. У тучных девочек с массой тела на 20—30% выше идеальной менархе наступает раньше, чем у девочек с нормальной массой тела. И наоборот, занятия некоторыми видами спорта или балетом, недоедание и хронические изнуряющие болезни обусловливают обычно задержку менархе.
Зрелый яичник
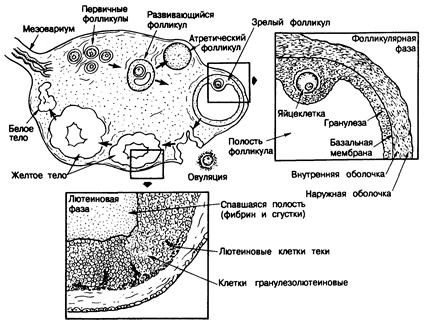
Морфология. Анатомическое строение и функция яичника у взрослых женщин схематически показаны на рис. 331-2. Под влиянием гонадотропинов активируется группа первичных фолликулов, и на 6—8-й день менструального цикла один из них становится зрелым, или «доминантным», т. е. в нем ускоряется рост клеток гранулезы и увеличивается заполненная жидкостью полость. Остальные активированные фолликулы начинают дегенерировать, что напоминает процесс атрезии, которому подвергаются другие фолликулы во время эмбриогенеза. Непосредственно перед овуляцией в яйцеклетке доминантного фолликула возобновляется мейоз, и первое мейотическое деление завершается формированием первого полярного тела. В результате накопления жидкости в фолликуле его полость быстро увеличивается и достигает размера 10—15 мм, поверхность становится тоньше, и образуется коническое выпячивание. Примерно через 16—23 ч после того, как уровень ЛГ достигает максимума, или через 24—36 ч после начала выброса ЛГ происходит овуляция из доминантного фолликула. Его стенка в области выпячивания разрывается, и яйцеклетка вместе с окружающими клетками гранулезы, называемыми клетками кумулюса, выталкивается. Разрыв объясняют действием гидролизующих ферментов на поверхность фолликула, возможно, с участием простагландинов. После оплодотворения яйцеклетки сперматозоидом начинается второе мейотическое деление, а затем образуется второе полярное тело. В проовулировавшем фолликуле начинает формироваться желтое тело; сохранившиеся клетки гранулезы и теки увеличиваются в размерах и накапливают липиды и желтый пигмент лютеин, становясь тем самым «лютеинизированными». Базальная мембрана, отделяющая клетки гранулезы от стромы и кровеносных сосудов, распадается, и капилляры, фибробласты и лимфатические протоки из теки внедряются в слой клеток гранулезы и достигают центральной полости, заполняя ее кровью. Через 14±2 дня (активная жизнь желтого тела) начинаются регрессия сосудов и атрофия желтого тела, завершающаяся его замещением фиброзным рубцом, т. е. образованием белого тела. Факторы, определяющие время существования желтого тела, неизвестны. Однако, если возникает беременность, желтое тело сохраняется под влиянием плацентарных или хориогонических гонадотропинов и вырабатывает прогестерон, который поддерживает беременность на ранних ее стадиях.
Рис. 331 -2. Изменения в зрелом яичнике в течение полного 28-дневного цикла.
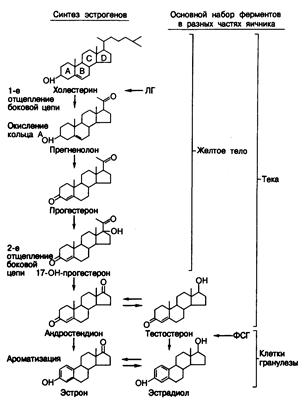
Образование гормонов. Стероидные гормоны. Как и другие стероидные гормоны, стероиды яичников образуются из холестерина (рис. 331-3). В яичниках холестерин может синтезироваться и de novo из двууглеродных предшественников; другим его источником служат липопротеиды низкой плотности (ЛПНП) крови. В обоих случаях он служит субстратом образования стероидных гормонов (рис. 331 -4). Считается, что практически все клетки яичника обладают полным набором ферментов, необходимых для превращения холестерина в эстрадиол (см. рис. 331-3). Однако в яичнике клетки разных типов содержат неодинаковое количество этих ферментов, и поэтому в отдельных участках органа преимущественно вырабатываются те или иные стероиды. Например, желтое тело в основном вырабатывает прогестерон и 17-гидроксипрогестерон, а клетки теки и стромы пре
Рис. 331-3. Основной путь биосинтеза стероидных гормонов в яичниках. Хотя любая клетка яичника содержит, вероятно, полный набор ферментов, требующихся для образования'эстрадиола из холестерина, количество отдельных ферментов и соответственно главные синтезируемые гормоны в клетках разного типа оказываются неодинаковыми. Основные наборы ферментов в клетках желтого тела, стромы и гранулезы ограничены скобками. В результате эти клетки вырабатывают преимущественно прогестерон и 17-ОН-прогестерон, андрогены и эстрогены соответственно. Важнейшие точки влияния ЛГ и ФСГ на процессы биосинтеза показаны горизонтальными стрелками.
вращают холестерин в андрогены — андростендион и тестостерон. Клетки гранулезы особенно активны в превращении андрогенов в эстрогены, и в качестве субстратов этого процесса используют андрогены, синтезируемые не только в них самих, но и в соседних клетках теки. На рис. 331-3 и 331-4 показаны и основные точки действия ЛГ и ФСГ. Например, ЛГ регулирует главным образом первый этап биосинтеза стероидных гормонов, а именно превращение холестерина в прегненолон, а также участвует в индукции ферментов, катализирующих последующие стадии этого процесса. ФСГ же регулирует конечный этап, т. е. ароматизацию андрогенов в эстрогены. Таким образом, ЛГ и без ФСГ способен усиливать поток субстратов и образование андрогенов и/или прогестерона, а ФСГ без ЛГ неэффективен, так как снижается количество субстратов для ароматизации. Эстрогены. Природные эстрогены — это 18-углеродные стероиды, характеризующиеся ароматическим кольцом А, фенольной гидроксильной группой в положении С-3 и либо гидроксильной (эстрадиол), либо кетоновой (эстрон) группой в положении С-17 (см. рис. 331-3). (Нумерацию атомов в кольцах стероидов см. на рис. 330-1.) Основным эстрогеном, секретируемым яичниками и наиболее активным из природных эстрогенов, является эстрадиол. Яичники секретируют и эстрон, но в основном он образуется вне железы в процессе превращения андростендиона в периферических тканях. В моче из эстрогенов в наибольшем количестве присутствует эстриол (16-гидроксиэстрадиол). Он образуется путем 16- гидроксилирования эстрона и эстрадиола. Гидроксилирование эстрогенов в положении С-2 или С-4 приводит к образованию катехол-эстрогенов, которые могут действовать как внутриклеточные медиаторы определенных эффектов эстрогенов. Эстрогены способствуют формированию вторичных половых признаков у женщин и вызывают рост матки, утолщение слизистой оболочки влагалища, разжижение слизи шейки матки и развитие протоков в молочных железах. Механизм действия эстрогенов в тканях-мишенях сходен с
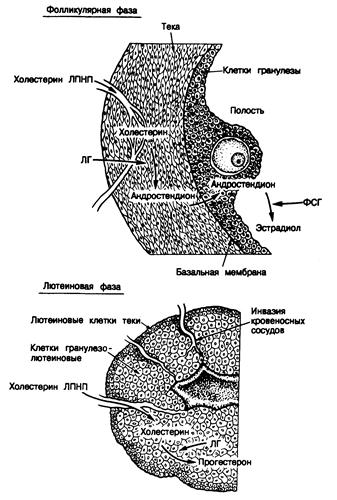
Рис 331 -4. Клеточные взаимодействия в яичнике в течение фолликулярной (вверху) и лютеиновой (внизу) фазы. Обозначения: ЛПНП — липопротеиды низкой плотности, ФСГ — фолликулостимулирующий гормон, ЛГ—лютеинизирующий гормон. (По В. R. Сагг, идр., 1982.)
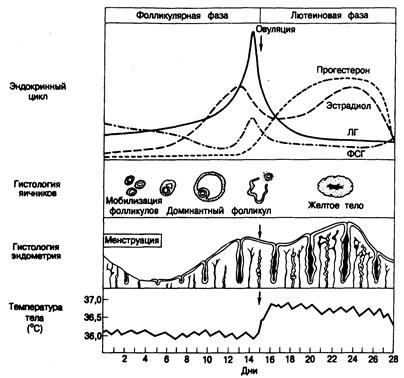
таковыми других стероидных гормонов и предполагает связывание со специфическим рецепторным белком цитозоля, последующее изменение конформации рецептора в транслокацию гормонрецепторного комплекса в ядро, где он присоединяется к ДНК и запускает транскрипцию информационных РНК. Последние в свою очередь обусловливают увеличение синтеза белка в цитоплазме клеток (см. гл. 320). Прогестерон. Прогестерон — 21 -углеродный стероид (см. рис. 331 -3) — основной гормон, секретируемый желтым телом; способствует сохранению беременности. Он индуцирует секреторную активность эндометрия подготовленной эстрогенами матки, обеспечивая тем самым возможность имплантации оплодотворенной яйцеклетки. Прогестерон вызывает в эндометрии и децидуальную реакцию. Кроме того, он вызывает сокращение матки, увеличение вязкости слизи шейки матки, развитие железистой ткани молочных желез и повышение базальной температуры тела (термогенный эффект). Андрогены. В яичниках, главным образом в клетках стромы и теки, синтезируются различные 19-углеродные стероиды, включая дегидроэпиандростерон, андростендион, тестостерон и дигидротестостерон. Основным 19-углеродным стероидом является андростендион (см. рис. 330-3). Некоторое количество его секретируется в плазму, а остальное превращается в эстрогены в клетках гранулезы или в тестостерон в интерстициальной ткани. Превращение андростендиона в тестостерон и эстрогены может происходить и на периферии. Истинными андрогенами, способными взаимодействовать с соответствующими рецепторами и обусловливать тем самым признаки вирилизации у женщин, являются только тестостерон и дигидротестостерон (см. гл. 46 и 330). Другие гормоны. Роль других гормонов, синтезируемых яичниками, в физиологии человека остается неясной. Релаксин _ полипептидный гормон, вырабатываемый у человека желтым телом и отпадающей оболочкой матки, вызывает у животных размягчение шейки матки и разрыхление лонного сочленения, подготавливая организм к родам. Фолликулярный иигибин, или фолликулостатин (эквивалент тестикулярного ингибина), секретируется фолликулами и, по-видимому, регулирует секрецию ФСГ гипоталамо-гипофизарным комплексом. Регуляторный белок фолликулов (РБФ) из фолликулярной жидкости человека тормозит секрецию и рост клеток гранулезы. Гонадокринины — пептиды, выделенные из фолликулярной жидкости крысы, стимулируют секрецию гипофизом ФСГ и ЛГ как in vitro, так и in vivo. Кроме того, в гонадах представителей обоих полов присутствует вещество, идуцирующее мейоз (ВИМ), которое запускает начальные стадии мейоза, что в яичниках происходит раньше, чем в яичках. В отличие от этого яички плода мужского пола секретируют преимущественно вещество, предотвращающее мейоз (ВПМ), которое препятствует инициации мейоза до начала полового созревания, когда преобладает образование ВИМ. Нормальный менструальный цикл. Менструальный цикл включает фолликулярную или пролиферативную, и лютеиновую, или секреторную, фазы (рис. 331-5). Секреция ФСГ иЛГ контролируется по механизму отрицательной обратной связи стероидами яичников (особенно эстрадиолом) и, вероятно, ингибином, но реакция гонадотропинов на различное содержание стероидов непостоянна. По мере увеличения уровня эстрогенов секреция ФСГ прогрессивно снижается — типичная отрицательная обратная связь. В отличие от этого секреция ЛГ максимально ингибируется малыми количествами эстрогенов, а при повышении их уровня и хронически повышенной концентрации эстрадиола — возрастает. Это так называемая регуляция по механизму положительной обратной связи. Действие эстрогенов по механизму отрицательной обратной связи замыкается как в гипоталамусе, так и в гипофизе, а по механизму положительной обратной связи — главным образом на уровне гипофиза. Продолжительность нормального менструального цикла определяют как время от начала одного менструального кровотечения до начала следующего. У женщин детородного возраста менструальный цикл длится в среднем 28±3 дня, а средняя продолжительность менструации 4±2 дня. В период менархе и перед менопаузой менструальные циклы более продолжительны. В конце каждого менструального цикла и при ослаблении функции желтого тела содержание эстрогенов и прогестерона г. плазме снижается, а уровень ФСГ в крови повышается. Под влиянием возрастающих количеств ФСГ начинается мобилизация фолликулов, что приводит к формированию доминантного в следующем цикле фолликула. После начала менструации фолликулы продолждают созревать, но уровень ФСГ снижается. Примерно за 8—10 дней до выброса ЛГ в середине цикла содержание эстрадиола в плазме начинает повышаться. Это происходит из-за его секреции клетками гранулезы растущего доминантного фолликула. Содержание ЛГ также начинает увеличиваться (положительная обратная связь). Перед овуляцией секреция эстрадиола достигает максимума, а затем снижается. Дальнейший подъем уровня ЛГ в плазме обусловливает созревание фолликула, вслед за чем происходят его разрыв и овуляция (через 16—23 ч после пика ЛГ). Одновременно с повышением уровня ЛГ незначительно увеличивается содержание ФСГ в плазме, физиологическое значение этого явления остается неясным. Приблизительно в середине цикла начинает возрастать концентрация прогестерона в плазме, что облегчает действие эстрадиола на секрецию ЛГ по механизму положительной обратной связи.
Рис. 331.5. Изменения гормональной среды, ткани яичников и эндометрия, а также базальной температуры тела и взаимосвязь этих изменений в ходе нормального менструального цикла.
В начале лютеиновой фазы содержание гонадотропинов в плазме снижается, а уровень прогестерона увеличивается. Вторичное повышение секреции эстрогенов способствует дальнейшему угнетению секреции гонадотропинов. К концу лютеиновой фазы уровень прогестерона и эстрогенов уменьшается, а содержание ФСГ возрастает, инициируя развитие следующего фолликула (обычно в другом яичнике) и следующий менструальный цикл. В ответ на меняющийся уровень гормонов яичников в плазме возникают выраженные изменения эндометриальной выстилки полости матки (см. рис. 331-5). Одновременно со снижением содержания эстрогенов и прогестерона в плазме и ослаблением функции желтого тела на поздних этапах лютеиновой фазы спиральные артериолы, снабжающие кровью эндометрий, резко сужаются, что приводит к ишемическому нейрозу, эсквамации эндометрия и кровотечению. Этот спазм сосудов обусловлен местно синтезирующимися простагландинами. Начало кровотечения — это первый день менструального цикла. До 4—5-го дня цикла эндометрий остается тонким. В пролиферативную фазу эстрогены вызывают рост желез эндометрия. Повышающееся после овуляции содержание прогестерона способствует дальнейшему утолщению эндометрия, но замедляет его быстрый рост. Затем эндометрий вступает в секреторную фазу, которая характеризуется извитостью желез, скрученными спиральными артериолами и секрецией. При ослаблении функции желтого тела (если не произошло зачатия) происходят изменения, приводящие к следующей менструации. Для овуляторного цикла характерны двухфазные изменения базальной температуры тела, обусловленные меняющимся содержанием прогестерона (см. рис. 331-5). Базальная температура тела после овуляции повышается на 0,3—0,5°С и сохраняется на этом уровне в течение всей лютеиновой фазы, а к началу следующей менструации восстанавливается исходная температура (36,2—36,4°С) (см. гл. 9). Клеточные взаимодействия в яичнике в ходе нормального цикла. Лютеинизирующий гормон стимулирует окружающие фолликул клетки теки к выработке андрогенов, и андростендион диффундирует через базальную мембрану фолликула в клетки гранулезы, где он ароматизируется в эстроген (см. рис. 331-3 и 331-4). Повышение уровня ФСГ в конце предыдущего менструального цикла стимулирует рост и мобилизацию первичных фолликулов, усиливая пролиферацию клеток гранулезы, что приводит к формированию доминантного фолликула. ФСГ повышает также активность и количество ферментов ароматизации в клетках гранулезы, превышающих андростендион в эстроген. Повышенная секреция эстрадиола вызывает увеличение числа эстрадиоловых рецепторов и дальнейшую пролиферацию клеток гранулезы. На поздних стадиях фолликулярной фазы ФСГ совместно с эстрадиолом индуцирует рецепторы ЛГ на клетках гранулезы. Действуя через эти рецепторы, ЛГ способствует увеличению секреции прогестерона в середине цикла. Считается, что количество продуцируемого фолликулом прогестерона лимитируется доступностью холестерина ЛПНП, служащего субстратом стероидогенеза, а также тем, что большая часть образуемого прогестерона подвергается в клетках теки дальнейшему метаболизму в андростендион. До овуляции клетки гранулезы фолликула погружены в фолликулярную жидкость, но плохо снабжаются циркулирующей кровью и, следовательно, ЛПНП плазмы. После овуляции клетки гранулезы васкуляризируются и приобретают способность использовать в качестве главного субстрата для синтеза прогестерона желтым телом холестерин ЛПНП плазмы (см. рис. 331-4). Таким образом, повышенный синтез желтым телом прогестерона обусловливается повышением доступности субстрата. Максимальная секреция прогестерона желтым телом происходит на 8-й день после овуляции, т. е. во время максимальной васкуляризации клеток гранулезы.
Менопауза
Менопауза — это фаза климактерического периода, наступающая после окончания последнего менструального кровотечения у женщин. В это время постепенно, но прогрессирующе снижается функция яичников и развиваются разнообразные эндокринные, соматические и психические изменения. У женщины менструальные кровотечения чаще прекращаются в 50—51 год. Поскольку ожидаемая продолжительность жизни женщин в настоящее время приближается к 80 годам, примерно треть жизни приходится на годы прекращения репродуктивной функции. Перед менопаузой характер менструальных циклов нарушается, интервалы между менструациями становятся короче из-за сокращения продолжительности фолликулярной фазы цикла. Кроме того, несмотря на сохранение овуляторных циклов, содержание ФСГ иЛГ в плазме увеличивается. Таким образом, перед менопаузой яичники, по-видимому, становятся менее чувствительными к гонадотропинам. Менопауза свидетельствует об истощении фолликулов яичников. Уменьшение числа яйцеклеток начинается еще во внутриутробном периоде, а ко времени наступления менопаузы остается лишь несколько яйцеклеток, которые, очевидно, лишены функции. В репродуктивном возрасте вследствие овуляции теряется лишь небольшое число яйцеклеток; большинство фолликулов и содержащихся в них яйцеклеток перестают функционировать в результате атрезии. Прекращение развития фолликулов приводит к снижению продукции эстрадиола и других гормонов, что в свою очередь обусловливает выпадение тормозных влияний на гипоталамо-гипофизарные центры по механизму отрицательной обратной связи. Вследствие этого возрастает содержание гонадотропинов в плазме, причем количество ФСГ увеличивается раньше и в большей степени, чем ЛГ (см. рис. 331-1; рис. 331-6). Повышение уровня ФСГ по сравнению с уровнем ЛГ у женщин в постклимактерическом периоде может произойти в результате снижения секреции ингибина яичниками, менее быстрого клиренса ФСГ, чем ЛГ, из плазмы из-за большего содержания в нем сиаловых кислот, а также утра ты механизма положительной обратной связи, предполагающего влияние эстрадиола на продукцию ЛГ. Внутривенное введение ЛГРГ женщинам в постклимактерическом периоде способствует усилению секреции как ФСГ, так и ЛГ, что характерно для повышенной секреторной активности гипоталамо-гипофизарной системы и при других формах первичной недостаточности яичников.
Рис. 331 -6. Различия в концентрации гормонов у женщин в репродуктивном возрасте и в период менопаузы. Обозначения: ФСГ—фолликулостимулирующий гормон, ЛГ—лютеинизирующий гормон, Е, — эстрадиол-17b, Е—эстрон, D4А — андростендион, Т — тестостерон. (По S. S. С. Yen a. R. В. Jaffe 1986, D. R. Mishell Jr. a. V. Davajan.)
В постклимактерический период яичники у женщин уменьшены в размерах, а сохранившиеся в них клетки имеют преимущественно стромальное происхождение. Содержание эстрогенов и андрогенов в плазме понижено, но они все же присутствуют в крови. До менопаузы андростендион плазмы почти в равных долях продуцируют надпочечники и яичники; после менопаузы яичники уже не вырабатывают андростендион, вследствие чего содержание его снижается на 50% (см. рис. 331 -6). Однако в менопаузе яичники продолжают секретировать тестостерон; по всей вероятности, эту функцию взяли на себя клетки стромы, Эстрогены крови у овулирующих женщин имеют два источника. В среднем 60% продукции эстрогенов во время менструального цикла приходится на эстрадиол, образуемый главным образом яичниками, а остальная часть — на эстрон, продуцируемый в основном внежелезистыми тканями из андростендиона. После менопаузы внежелезистое образование эстрогенов становится главным источником их синтеза. Яичники в менопаузе продуцируют лишь минимальное количество эстрогенов, и оофорэктомия в этом периоде не сопровождается в дальнейшем сколько-нибудь заметным снижением их уровня. Содержание в плазме эстрадиола, главного эстрогена, секретируемого фолликулами, у женщин в постклимактерическом периоде ниже, чем содержание эстрона. Скорость периферического образования эстрона в период менопаузы у женщин несколько возрастает, так что продукция эстрона оказывается лишь ненамного меньшей, чем до менопаузы, несмотря на понижение уровня андростендиона в плазме. Поскольку основным местом внежелезистого образования эстрогенов служит жировая ткань, у тучных женщин в постклимактерическом периоде периферическое содержание эстрогенов может даже увеличиваться, так что общая скорость продукции этих соединений будет такой же или даже большей, чем у женщин в предклимактерическом периоде. Однако преимущественно будет образовываться эстрон, а не эстрадиол. К наиболее частым симптомам менопаузы относятся вазомоторная неустойчивость (горячие приливы), атрофии эпителия мочеполовых путей и кожи, уменьшение размеров грудных желез и остеопороз. Примерно у 40% женщин в постклимактерическом периоде симптомы настолько выражены, что заставляют прибегать к помощи врача. Патогенез горячих приливов неясен. Существует тесная связь во времени между началом приливов и секреторными импульсами ЛГ. Определенную роль в возникновении этого феномена могут играть также изменения метаболизма катехоламинов, простагландинов, эндорфинов или нейротензинов наряду с низкой продукцией эстрогенов. Горячие приливы часто сопровождаются нервозностью, чувством тревоги, раздражительностью и депрессией; их связь с дефицитом эстрогенов не доказана. Уменьшение размеров органов женских половых путей и грудных желез во время менопаузы — это следствие недостатка эстрогенов. Эндометрий становится тонким и атрофичным (хотя у 20% женщин в постклимактерическом периоде регистрируют кистозную гиперплазию); аналогичным изменениям подвергается слизистая оболочка влагалища и мочеиспускательного канала. Существует тесная взаимосвязь между снижением уровня эстрогенов и развитием остеопороза. Остеопороз — это страшное бедствие, сопровождающее старение. Примерно у 20% женщин и 10% мужчин в возрасте между 60 и 90 годами возникают переломы позвонка или бедренных костей, причем наиболее часто их наблюдают у пожилых женщин белой расы. Такие переломы служат основной причиной смертности и инвалидности, и связанная с переломами смертность колеблется от менее 10% в возрастной группе 60—64 года до 30% и более среди лиц старше 80 лет. На развитие остеопороза влияет множество факторов, включая диету, физическую активность, курение и общее состояние здоровья, но особую роль в этом играет отсутствие эстрогенов. Женщины белой расы в постклимактерическом периоде более предрасположены к остеопорозу и его последствиям, поскольку у них и до менопаузы меньше плотность кости, а дальнейшее уменьшение этой плотности чревато для данной группы более тяжелыми последствиями. Еще одним доказательством того, что остеопороз — это болезнь отсутствия эстрогенов, служит более раннее его развитие у женщин с преждевременной менопаузой, обусловленной либо естественными причинами, либо хирургической кастрацией.
Дата добавления: 2015-02-02 | Просмотры: 1695 | Нарушение авторских прав |